免疫细胞膜分子种类繁多,分类的方法各有不同的角度。如前所述,CD抗原主要是应用单克隆抗体等技术,对于执行各种功能的细胞膜分子所进行的分类,几乎是包罗万象。在CD中,实际上包括了粘附分子中部分Ig超家族成员、大部分粘合素超家族成员和所有选择素成员。除此之外,从功能分类的角度,细胞膜分子还包括免疫活性细胞抗原识别受体、主要组织兼容性抗原、细胞因子受体、T和B淋巴细胞的分化抗原,这些内容将在本书有关的章节中分别阐述。本节主要介绍促分裂原及其在淋巴细胞上相应的结合分子以及各种IgFe受体。
一、促有丝分裂原受体
有丝分裂原(mitogen)来自植物蛋白或细菌产物,它能与多种细胞膜糖类及寡糖基分子结合,后者为促使丝分裂原受体、结合后能促使细胞活化和诱导细胞分裂。由于细胞膜含有不同糖基,故可认为是多克隆活化剂,它与抗原特异性的克隆活化是不同的。
T和B细胞表面都有分裂素受体,在体外可非特异性刺激静止淋巴细胞向母细胞转化。这种转化细胞DNA合成增加,出现细胞体积增大,胞浆增多,嗜碱性以及产生有丝分裂等形态变化。这种转化过程对研究淋巴细胞功能变化及淋巴细胞早期活化过程的生化变化甚为重要。这种淋巴细胞转化过程可用3H-TdR掺入,检测DNA合成变化或借光细胞转化,常用于T细胞功能检测。美洲商陆(PWM)可诱导T和B细胞转化,而细菌脂多糖(LPS)能引起B细胞转化(珍6-8)。
表6-8 常见促有丝分裂原的特征
| 名称 | 来源 | 分子量(kD) | 糖特异性 | 特性及应用 |
| 刀豆素A(ConA) | 刀豆 | 102 | α-D-mannosyl-
α-D-glucose | T细胞促有丝分裂原,分离细胞膜糖蛋白 |
| 植物血凝素(PHA) | 云豆 | 120 | N-actyl-D-galactosamine | T细胞促有丝分裂原 |
| 美洲商陆丝裂原(PNA) | 美洲商陆 | 32 | β-N-acetyl-D-glucosamine | B细胞、T细胞有丝裂原 |
| 大豆凝集素(SBA) | 花生 | 110 | D-galactosyl-(β1-3)-N-acetyl-D-galactosamine | 分离胸腺细胞亚群(皮质不成熟胸腺细胞与PNA凝集为PNA+细胞) |
| 大豆凝集素(SBA) | 大豆 | 120 | N-acetyl-D-galactosamine
D-galactose | 凝集免疫活性B细胞、净化人骨髓用于骨髓移植 |
二、Ig Fe受体
Ig分为IgM、IgD、IgA、IgD和IgE五类。各类Ig的不同功能主要与其结构有关。机体内许多细胞表面具有不同类IgFe的受体,通过Fe受体与IgFe的结合,参与Ig介导的生理功能或病理损伤过程。目前已鉴定明确的Fe受体有FeγR、FcαR和FcεR.
(一)FeγR
1.FeγR的结构和分布 FeγR可分为FeγRⅠ、FeγRⅡ和FeγRⅢ三类,它们的结构和分布有所不同。
(1)FeγRⅠ(CD64):为70kD穿膜糖蛋白,属Ig超家族,胞膜外区有3个C2结构,FeγRⅠ主要分布于单核细胞、巨噬细胞、中性粒细胞等。IFN-γ可刺激单核细胞、巨噬细胞中性细胞表达FeγRⅠ水平增加5~10倍,G-CSF也这种促进作用。
(2)FeγRⅡ(CD32),为40kD穿膜糖蛋白,属于Ig超家族,胞膜外区有2个C2结构,FeγRⅡ表达于除红细胞外的其它血细胞。
(3)FeγRⅢ(CD16):为50~70kD糖蛋白,属Ig超家族,有2个C2结构。主要分布于巨噬细胞、NK细胞和嗜酸性粒细胞,中性粒细胞表面。
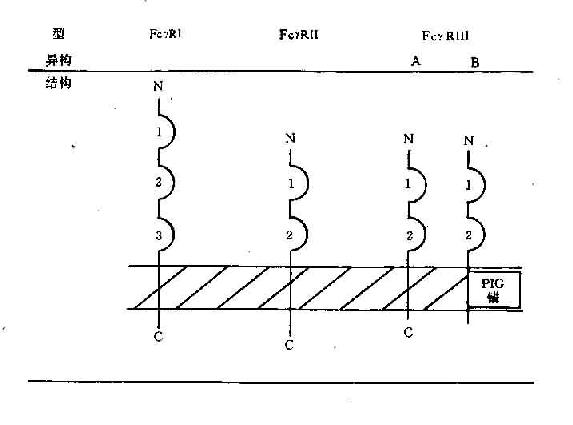
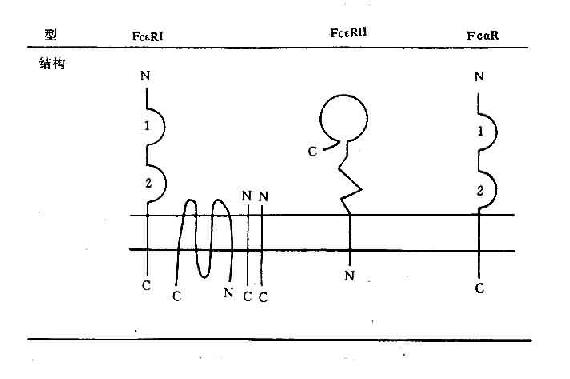
图6-2 人Feγ(上图)和人Feα、Feε受体(下图)结构示意图
2.FeγR的功能 FeγR的功能主要是通过髓样细胞和NK细胞来发挥的。
(1)单核-巨噬细胞:FeγRⅠ、Ⅱ和Ⅲ均可介导入单核细ADCC来杀伤肿瘤等靶细胞。这种ADCC效应为Mg2+依赖,并需要LFA-1等粘附分子参与。IFN-γ可促进单核细胞FeγRⅠ介导的杀伤作用.单核-吞噬细胞可能通过FeγRⅠ、Ⅱ、Ⅲ发挥调理吞噬和清除免疫复合物作用。
(2)中性粒细胞:新鲜分离的中性粒细胞不能通过FeγR溶解靶细胞,但在IFN-γ刺激后可通过FcγRⅠ和FcγRⅡ介导杀伤作用,对FcγRⅠ主要是诱导其表达水平升高,而对FcγRⅡ表达水平并未见改变,可能是通过杀伤机制的调节。GM-CSF也能通过FcγRⅡ明显增强中性细胞的杀伤水平。活化中性粒细胞通过FcγRⅠ、Ⅱ发挥调理吞噬和清除免疫复合物作用。
(3)嗜酸性粒细胞:未刺激的嗜酸性粒细胞没有杀伤作用,GM-CSF、TNF和IL-5等是嗜酸性粒细胞发挥ADCC效应的有效激活剂,在抗寄生虫和抗肿瘤中有重要作用。GM-CSF激活作用主要是通过FcγRⅡ介导的。
(4)NK细胞:通过FcγRⅢA介导ADCC杀伤肿瘤细胞等靶细胞,IL-2和IFN-γ可明显提高NK细胞的杀伤活性,但并不明显改变FcγRⅢ的表达水平。
(二)FcαR
FcαR(CD89)为分子量60kD的穿膜糖蛋白,属Ig超家族,胞膜外有2个C2功能区,呈中等亲和力,主要表达于单核细胞、巨噬细胞、中性粒细胞等,介导吞噬,ADCC以及炎症介质的释放。
(三)FcεR
FcεR的结构和分布 FcεR可分为FcεRⅠ和FcεRⅡ两类,它们的结构、分布以及介导的作用有所不同。
(1)FcεRⅠ:为高亲和力受体,K值10-9~10-10M。由α、β、γ-γ四条链组成。FcεRⅠ主要分布于嗜碱性粒细胞和肥大细胞。
(2)FcεRⅡ(CD23):低亲和力受体,分子量45kD,单链穿膜糖蛋白。胞膜外C端侧25kD部分又称为IgE结合因子(IgE-BF),FcεRⅡ可在蛋白水解酶裂解后形成可溶性CD23分子(Scd23)即IgE-BF。
2.FcεR的功能
(1)FcεRⅠ:嗜碱性粒细胞和肥大细胞具有高亲和力FcεRⅠ,当相应变应原与嗜碱性粒细胞、肥大细胞表面lgE/FcεRⅠ复合物结合后,使细胞脱颗粒,合成和释放多种介质,介导Ⅰ型速发型超敏反应。
(2)FcεRⅡ:FcεRⅡ为B细胞分化激活抗原,在变态反应性疾病患者PBMC中CD23密度明显增加,血清sCD23升高。sCD23具有BCGF活性,促进B细胞产生IgE,并与IL-4有协同作用。此外,FcεRⅡ还可介导IgE依赖的ADCC和容噬作用。
三、细胞因子受体
免疫细胞表面表达多种细胞因子受体(cytokine receptor),不同免疫细胞表达细胞因子受体的种类、密度和亲和力有所差别。
1.T细胞表面的细胞因子受体多种细胞因子可调节T细胞的功能,这是因为T细胞存在着相应的细胞因子受体,如IL-1R、IL-2R、IL-3R、IL-4R、IL-6R、IL-7R、IL-8R、IL-9R、IL-12R、TNF-αR、G-CSFR和TGF-βR等。静止和活化T细胞因子受体的数目和亲和力可有很大差别,如静止T细胞只表达IL-β链,与IL-2配体结合是中亲和力,当T细胞活化后同时表达IL-2Rα链和β链,并组成高亲和力受体。
2.B细胞表面的细胞因子受体多种细胞因子调节B细胞的活化、增殖和分化是通过与B细胞表面相应的细胞因子受体结合而发挥调节作用的。B细胞的细胞因子受体主要有IL-1R、IR-2R、IL-4R、IL-5R、IL-6R、IL-7R、IL-11R、IL-12R、IFN-γR、TNF-α和TGF-βR等。
3.单核-巨噬细胞表面的细胞因子受体单核-巨噬细胞表面有多种细胞因子受体,如MCFR、MIFR、MAFR、IFNR、M-CSFR、GM-CSFR等,相应的细胞因子作用后可调节单核-巨噬细胞的吞噬功能、细胞毒作用、MHCⅡ类抗原的表达以及单核因子的合成和释放。
四、羊红细胞受体
人T细胞表面有能与绵羊红细胞结合的受体,称为E受体,E受体在体外可与绵羊红细胞表面的配体在一定条件下结合,可在T细胞表面形成花环状,称为E花环。用这个方法可以检测人外周血T细胞的相对百分数,可作为判断人T细胞免疫功能指标之一。
E受体已证明是分子量为30~60kD的糖蛋白分子。应用单克隆抗体鉴定E受体,认为它是一种新的人T细胞分化抗原,命名为白细胞分化抗原2(Leu-5,OKT11,CD2)。目前认为E受体可能是介导Ca2+依赖的T细胞旁路活化途径。这一途径是与T细胞抗原受体活化途径完全不同的另一活化途径。
五、补体受体
Uhr等在60年代发现由抗原、抗体与补体分子形成免疫复合物与部分豚鼠淋巴细胞结合,如从复合物中除去补体,则复合物与细胞的结合明显减少。证明了细胞表面有补体受体(complement receptor,CR)的存在。其后应用抗绵羊红细胞抗体(A)和补体(C)致敏的绵羊红细胞(E)形成的复合物(EAC)可与细胞表面的补体结合形成花环,称为EAC花环。用以检测CR。
多数B细胞表面有CR,除B细胞外,单核 -巨噬细胞、中性粒细胞、K细胞、哺乳类动物血小板、人及灵长类红细胞上均有补体受体。B细胞的补体受体与其活化有关,补体受体也能促进吞噬细胞的吞噬作用。此外,如免疫粒附作用以及抗体依赖的细胞介导的细胞毒作用也与补体有关。
六、内分泌激素、神经递质和神经肽胺体
免疫细胞表面可具有多种激素、神经递质和神经肽的受体,如雌激素、甲状腺素、肾上腺皮质激素、肾上腺素、前列腺素E、生长激素、胰岛素等激素的受体,内啡肽、脑啡肽、P物质等神经肽受体,组胺、乙酰胆碱、5-羟色胺、多巴胺等神经递质受体。免疫细胞表面的激素、神经肽和神经递质受体是机体神经内分泌免疫网络中的一个重要环节。