1975年著名免疫学家Burnet提出克隆选择学说,并以克隆清除(clonaldeletion)学说解释免疫耐受现象。他的观点曾对免疫学的发展产生深远影响。随着近代基础免疫学,尤其是免疫调节研究的迅速发展,当前对免疫耐受机制的认识已远远超越了这一学说当时的涵义。它的发生涉及到免疫应答过程中任何一个正、负调节系统。下述几种重要观点,其各自均有相应的实验证据。
一、克隆清除
Burnet的克隆选择学说提出体内约存在着102-107具有免疫活性的细胞克隆,每一克隆细胞都具有其特异的、能与其相应抗原决定簇起反应的受体。但处于未成熟阶段的T、B反应细胞系因接触抗原而被清除,则造成免疫耐受。现知大量未成熟自身反应性T细胞在胸腺内因接触相应的自身抗原后,发生程序性死亡而被清除,这是维持自身耐受最有效的机制。
克隆清除学说强调了免疫耐受诱导过程的中枢衰竭机制。这一学说尚不能解释许多客观存在的现象:①已建立的耐受性可以、甚至易于被破坏,即对原先的耐受原重新出现免疫应答;②给已建立耐受性的动物输注同品系正常动物的淋巴细胞(含反应克隆)并不能使动物恢复对耐受原的免疫应答;③将已建立耐受动物的淋巴细胞转移到同品系正常动物体内,能使其产生对相同抗原的耐受性;④成年机体对大多数自身抗原虽呈免疫耐受,但业已发现成年机体内可检测到对自身抗原起反应的T和B细胞克隆的存在。
以上事实提示,在免疫耐受,尤其是获得性免疫耐受的机体内,自身反应细胞克隆并未被彻底清除,而是处于功能受抑或无能状态(clonalanergy)。凡是细胞表面抗原受体被封闭,抗原不能与细胞表面受体结合,缺少T细胞或巨噬细胞的辅助作用,以及T细胞与巨噬细胞主动抑制作用,抗独特型网络的主动抑制等都可能参与耐受的诱导过程。导致反应细胞克隆的不应答,而不能发生正相免疫应答。
二、克隆不应答
(一)免疫活性细胞缺乏激活信号
现已知T细胞必须的激活信号至少包括:①由特异抗原与自身MHCⅠ类或Ⅱ类抗原的复合物激发的信号;②由协同刺激因子(costimulator)激发的信号。缺乏足够的激活信号则导致免疫不应答。目前认为,一些针对胸腺内不表达的自身抗原(如器官特异抗原)的自身反应性T细胞克隆存在于正常机体,但因带有这些自身抗原的细胞表面通常有具有MHCⅡ类抗原,因此不能激活相应T细胞克隆。
(二)免疫活性细胞激活受阻
1.免疫活性细胞表面抗原受体被封闭则可产生不应答。适量双价或多价抗原与免疫活性细胞表面抗原受体结合,受体聚集成帽状,使细胞活化而产生免疫应答(图13-1B)。而单价抗原(monomeric antigen)与免疫活性细胞表面抗原各个受体结合(图13-1A),抗原占据整个细胞的表面受体,对受体起封闭作用,则不能激活免疫细胞。如分子表面有许多相同重复决定簇的非胸腺依赖抗原在体内不易被分解,能与B细胞表面的抗原受体呈牢固、广泛交联,可使受体封闭。高剂量多价抗原使细胞表面抗原受体广泛交联,使液态镶嵌的细胞膜不能流动,膜受体呈“冻结”状态,细胞不被活化(图13-1C)。
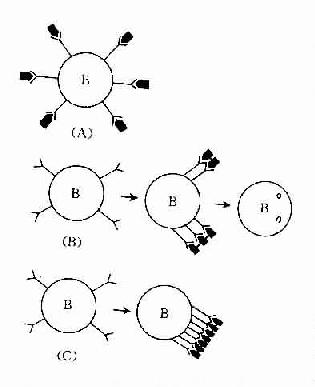
图13-1 B细胞表面抗原受体的封闭
(A)单价抗原占据B细胞表面抗原受体;
(B)适量双价抗原使B细胞表面受体交联成帽状,内吞;
(C)大量 多价抗原使B细胞表面受体交联“冻结”
2.抗原不能抵达免疫活性细胞表面有时机体在初次接触抗原后,产生抗体过剩,抗体与再次进入的抗原在体液中结合,使抗原不能到达细胞表面受体上,因而也可造成免疫无反应性。
(三)缺乏辅助细胞
胸腺依赖抗原(自然界大多数抗原均属此类)激发免疫应答均需TH细胞巨噬细胞的参与,若缺乏辅助细胞,免疫活性细胞单独不能作出有效应答。
1.缺乏辅助性T细胞(TH) 前已述及T/B细胞对同一抗原产生耐受性时,表现不同的特征。小剂量抗原便足以使T细胞产生耐受,此时B细胞虽未产生耐受,但因失去T细胞的辅助而不能活化,但是T细胞的耐受性维持时间大大超过B细胞(如图13-2所示)。B细胞虽恢复免疫应答,但T细胞仍处于耐受状态,因而出现T、B反应性呈分离状态的区域(表13-3)。这时,B细胞仍缺乏必要的T细胞辅助而不能产生有效的免疫应答。
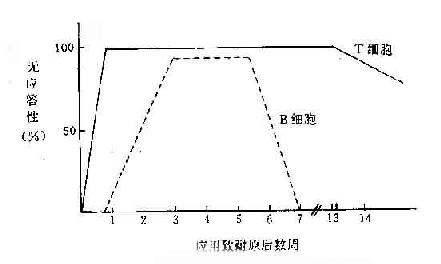
图13-2 T、B 细胞发生耐受的不同特征
2.缺乏巨噬细胞辅助巨噬细胞在免疫应答形成中,起着重要的摄取抗原、加工和呈递抗原的作用,从而参与了特异性免疫应答。所以巨噬细胞的功能缺陷也是耐受诱导的重要的原因。
表13-3 T细胞与B细胞耐受特征
| T细胞 | B细胞 | |
| 克隆清除 | 蛋白质抗原胸腺内诱导
机制:不成熟T细胞与抗原高亲和力结合导致和序性细胞死亡(PCD) | 蛋白质抗原及非蛋白质抗原导部位骨髓或周围尚不肯定
机制、:程序性细胞死亡(PCD) |
| 克降不应答 | APC缺乏协同刺激因子 | 多价抗原与不成熟B细胞结合 |
| 抗原性质 | 可溶性蛋白质静脉或口服无佐剂参与 | 大剂量多糖类抗原多聚蛋白质抗原(重复表位)无TH参与 |
| 耐受期限 | 期限长 | 期限短 |
| 抗原耐受剂量 | 剂量低 | 剂量高 |
三、抑制细胞的作用
(一)TS细胞的作用
70年代Gershon等首先提出TS细胞的现象。将耐受动物的脾或胸腺细胞转输给同品系正常动物后,使后者获得耐受性,又称为传染性耐受(infectious tolerance)。如果在转输前将脾细胞用抗Thy-1血清加补体处理,去除T细胞,则受体动物不会发生过继性耐受。
TS细胞的作用通常是抗原特异性的,它可能是通过阻止抗原呈递,阻断TH细胞的功能。抑制B细胞分化以及阻断B细胞分化为抗体分泌细胞等环节发挥作用。
(二)自然抑制细胞的作用
自然抑制(natural suppressor,NS)细胞主要抑制T细胞参与的免疫应答,无抗原特异性。这些细胞可能在新生与成年动物的耐受诱导中均起作用。NS细胞形态上为大颗粒淋巴细胞(large granularlymphocytes,LGL),见于胚胎及新生期、出生后数天内消失,抗原不能诱导,表面无T、B细胞特有的标志、对B细胞无抑制作用,成年动物照射后,先是NS细胞的再现,照射后短期内导入抗原,继之便出现TS细胞。照射促使骨髓移植物存活,可能与此种机制有关。抑制功能:混合淋巴细胞反应、TC细胞生成GVH反应。
(三)巨噬细胞的抑制作用
已有文现献报告抑制性单核-巨噬细胞亚群的存在。其抑制作用可能是由花生四烯酸代谢产物所介导。因阿司匹林和消炎痛能逆抑制作用。我国学者发现耐受动物的腹腔巨噬细胞有抑制同系正常动物混合淋巴细胞反应的作用,此作用并有抗原特异性。在对照组中,正常动物的巨噬细胞却能增强抗原特异性的混合淋巴细胞反应。
(四)抗独特型网络的作用
每个T、B细胞克隆均具有其独特型。B细胞表面及分泌的免疫蛋白超变区抗原结合部位是独特型的物质基础。免疫球蛋白独特型结构本身具抗原性,被相应细胞克隆识别而产生抗独特型抗体可进一步诱导抗-抗独特型第一系列连锁反应。对免疫应答起“自限”作用。T、B细胞参与的免疫应答均受独特特型网络的调节。有人报道给新生鼠注射抗特型抗体导致长期独型耐受,而对成年动物注射抗独特型抗体,可引起短暂的独特型耐受。
抗独特抗体所引起的耐受性,仅针对抗体的独特型部分,机体对抗原其他决定簇的免疫应答依然存在。