目前认为上述诱因导致自身免疫的形成是通过多种机制而实现的。不同情况下由不同机制起主要作用,有时也可能由几种机制同时或先后起作用。
一、禁忌株突变
通过体细胞突变可能产生被Burnet称为禁忌的细胞株。使在正常情况下已遭克隆清除的自身反应性T.B细胞再现。
二、T、B细胞活化信号的出现
正常人体内存在一些自身反应性T.B细胞克隆,因缺乏足够的激活信号而处于无应答状态(失活或静止状态)。一旦获得足够的激活信号,便可产生正相应答。
现有实验证明,在某些自身免疫病中受累器官细胞表面,或是出现了原先不表达的DR/I-A抗原,或是DR/IA的抗原较原先表达增多。病毒感染导致自身免疫病的可能机制是通过机体释放的淋巴因子,如IFN-r和IL-1等。使得原先不表达MHC-Ⅱ类抗原的组织细胞出现DR/I-A抗原。同种免疫可以诱导出自身免疫,可能亦与此机制有关。例如造成GVH病动物体内角质细胞表面表达I-A抗原,I-A阳性T细胞增加。这些质和量改变的MHCⅡ类抗原可激发自身免疫反应。实验证明,注射抗H-21区抗原的抗体可预防或减弱小鼠实验性重症肌无力、脑脊髓和Ⅱ型胶原性关节炎等。
三、B细胞被多克隆激活
在T细胞处于耐受的情况下,用B细胞多克隆激活剂直接刺激B细胞,可导致自身抗体的产生。例如,给小鼠注射LPS与自身抗原并用,则可导致特异性自身抗体生成。甲状腺球蛋白与LPS合用可诱发抗甲状腺球蛋白特异抗体。
四、TH细胞旁路激活
此学说的基本观点是机体对某些自身抗原的耐受性仅是由于T细胞处于耐受状态,使B细胞缺少辅助信号而不能有效地活化。一旦通过旁路机制获取T细胞辅助信号,B细胞即可活化,产生自身抗体(图16-1,2)
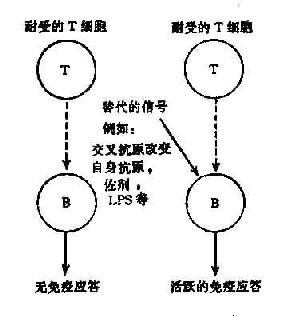
图16-1 T细胞旁路活化机制
图16-2 破坏T细胞对自身抗原的耐受性
佐剂、病毒、细胞产物、以及某些药物具有非特异地激活T、B细胞的作用(多克隆激活),它们诱发自身免疫的机制可能就是通过替代因子的旁路作用,绕过耐受的T细胞,使B细胞活化。
T细胞旁路机制不但可诱导自身抗体生成,也能引起细胞介导的自身免疫性损伤,如实验性变态反应性脑脊髓炎。甲状腺炎等。
五、自身反应克隆脱抑制
TS细胞能抑制自身反应细胞的激活,TS细胞数量或功能降低,TH和(或)TCS细胞数量增多或活跃,使自身反应细胞发生脱抑制而功能亢进,都可导致自身免疫的发生。
早期系统的实验资料曾对SLE样综合征自发品系NZB及NAB/NZW小鼠自出生后逐月进行免疫功能检测。从时间顺看,这种动物体内最早出现的是T细胞调节功能紊乱,约在出生后1个月,动物血清中胸腺激素活性下降,TS细胞功能减退。T细胞对诱导免疫耐受呈抵抗,然后再出现B细胞功能亢进。接近3月龄时,血清中才出现抗核抗体、抗淋巴细胞表面抗体及其他抗血细胞抗体等多种自身抗体。
受动物实验启示,临床免疫工作者测定各种自身免疫病患者外周血T细胞亚群变化,也发现类似变化。应用抗T细胞分化抗原单克隆抗体测定时,发现SLE和类风湿关节炎患者在病性活动期,TS细胞比例下降,用Con A在体外刺激病人淋巴细胞不诱发非特异的TS细胞。
六、独特型网络激活
独特型-抗独特型的相互作用极为复杂。在不同的条件下,对自身免疫产生不同的调节作用,有时抑制(自身耐受),有时增强。外来或体自身抗原均可通过此网络活化而造成自身免疫。根据独特型网络的基本理论,有以下几种可能性(图16-3):
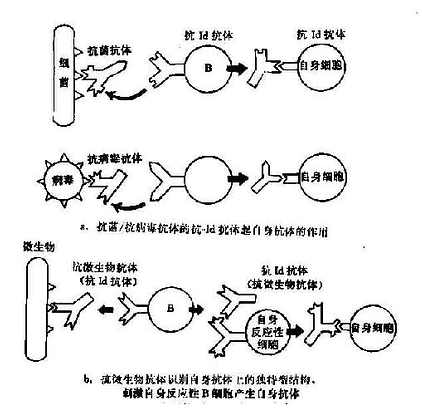
图16-3 独特型相互作用形成自身免疫
1.针对抗原(细菌、病毒、激素等)所产生的抗体,具有与自身细胞膜抗原相似或相同的决定簇,故其所激发的抗独特型抗体能与自身细胞表面抗原结事(图16-3a)。已有实验证据揭示这种可能性的存在,如小鼠注射胰岛素后所产生的抗体能进一步诱发生成抗独特型抗体。这种抗体不仅能与激发抗体结合,也能与胰岛素受体结合,造成胰岛素抵抗型糖尿病。
2.抗微生物抗体本身可能就是一种抗自身抗体的抗独特型抗体,它能激活具有独特表位的自身的反应性B细胞(图16-3b)。