大多数骨骼肌(skeletal muscle)借肌健附着在骨骼上。分布于躯干和四肢的每块肌肉均由许多平行排列的骨骼肌纤维组成,它们的周围包裹着结缔组织。包在整块肌外面的结缔组织为肌外膜(epimysium),它是一层致密结缔组织膜,含有血管和神经。肌外膜的结缔组织以及血管和神经的分支伸入肌内,分隔和包围大小不等的肌束,形成肌束膜(perimysium)。分布在每条肌纤维周围的少量结缔组织为肌内膜(endomysium),肌内膜含有丰富的毛细血管(图6-1)。各层结缔组织膜除有支持、连接、营养和保护肌组织的作用外,对单条肌纤维的活动、乃至对肌束和整块肌肉的肌纤维群体活动也起着调整作用。
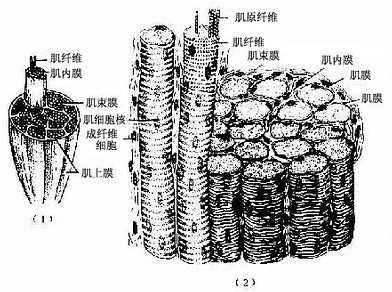
图6-1 骨骼肌与周围结缔组织膜
(1)一块骨骼肌模式图,示肌外膜、肌束膜和肌内膜
(2)骨骼肌纤维纵横切面
(一)骨骼肌纤维的光镜结构
骨骼肌纤维为长柱形的多核细胞(图6-1),长1~40mm,直径10~100μm。肌膜的外面有基膜紧密贴附。一条肌纤维内含有几十个甚至几百个细胞核,位于肌浆的周边即肌膜下方。核呈扁椭圆形,异染色质较少,染色较浅。肌浆内含许多与细胞长轴平行排列的肌原纤维,在骨骼肌纤维的横切面上,肌原纤维呈点状,聚集为许多小区,称孔海姆区(Cohnheim field)。肌原纤维之间含有大量线粒体、糖原以及少量脂滴,肌浆内还含有肌红蛋白。在骨骼肌纤维与基膜之间有一种扁平有突起的细胞,称肌卫星细胞(muscle satellite cell),排列在肌纤维的表面,当肌纤维受损伤后,此种细胞可分化形成肌纤维。
肌原纤维(myofibril)呈细丝状,直径1~2μm,沿肌纤维长轴平行排列,每条肌原纤维上都有明暗相间、重复排列的横纹(cross striation)。由于各条肌原纤维的明暗横纹都相应地排列在同一平面上,因此肌纤维呈现出规则的明暗交替的横纹。横纹由明带和暗带组成(图6-2)。在偏光显微镜下,明带(light band)呈单折光,为各向同性(isotropic),又称I带;暗带(dark band)呈双折光,为各向异性(anisotropic),又称A带。在电镜下,暗带中央有一条浅色窄带称H带,H带中央还有一条深 M线。明带中央则有一条深色的细线称Z线。两条相邻Z线之间的一段肌原纤维称为肌节(sarcomere)。每个肌节都由1/2I带+A带+1/2I带所组成(图6-3,6-4)。肌节长约2~2.5μm,它是骨骼肌收缩的基本结构单位。因此,肌原纤维就是由许多肌节连续排列构成的。
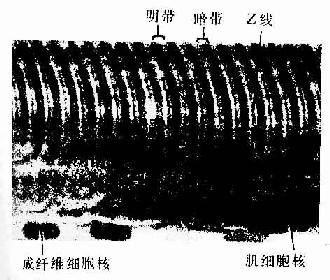
图6-2 人骨骼肌纤维纵切面

图6-3 兔骨骼肌纤维纵切
Z Z线 (白求恩医科大学尹昕、朱秀雄教授供图)
(二)骨骼肌纤维的超威结构
1.肌原纤维 肌原纤维是由上千条粗、细两种肌丝有规律地平行排列组成的,明、暗带就是这两种肌丝排布的结果(图6-4)。粗肌丝(thick filament)长约1.5μm,直径约15nm,位于肌节的A带。粗肌丝中央借M线固定,两端游离。细肌丝(thin filathent)长约1μm,直径约5nm,它的一端固定在Z线上,另一端插入粗肌丝之间,止于H带外侧。因此,I带内只有细肌丝,A带中央的H带内只有粗肌丝,而H带两侧的A带内既有粗肌丝又有细肌丝(图6-4);所以在此处的横切面上可见一条粗肌丝周围有6条细肌丝;而一条细肌丝周围有3条粗肌丝(图6-4)。两种肌丝肌在肌节内的这种规则排列以及它们的分子结构,是肌纤维收缩功能的主要基础。
粗肌丝的分子结构:粗肌丝是由许多肌球蛋白分子有序排列组成的。肌球蛋白(myosin)形如豆芽,分为头和杆两部分,头部如同两个豆瓣,杆部如同豆茎。在头和杆的连接点及杆上有两处类似关节,可以屈动。M线两侧的肌球蛋白对称排列,杆部均朝向粗肌丝的中段,头部则朝向粗肌丝的两端的两端并露出表面,称为横桥(cross bridge)(图6-4)。M线两侧的粗肌丝只有肌球蛋白杆部而没有头部,所以表面光滑。肌球蛋白头部是一种ATP酶,能与ATP结合。只有当肌球蛋白分子头部与肌动蛋白接触时,ATP酶才被激活,于是分解ATP放出能量,使横桥发生屈伸运动。
细肌丝的分子结构:细肌丝由三种蛋白质分子组成,即肌动蛋白、原肌球蛋白和肌原蛋白。后二种属于调节蛋白,在肌收缩中起调节作用。肌动蛋白(actin)分子单体为球形,许多单体相互接连成串珠状的纤维形,肌动蛋白就是由两条纤维形肌动蛋白缠绕形成的双股螺旋链。每个球形肌动蛋白单体上都有一个可以与肌球蛋白头部相结合的位点。原肌球蛋白(tropomyosin)是由较短的双股螺旋多肽链组成,首尾相连,嵌于肌动蛋白双股螺旋链的浅沟内。肌原蛋白(troponin)由3个球形亚单位组成,分别简称为TnT、 TnI和 Tnc 。肌原蛋白借TnT而附于原肌球蛋白分子上, TnI是抑制肌动蛋白和肌球蛋白相互作用的亚单位, Tnc 则是能与Ca2+相结合的亚单位(图6-4)。
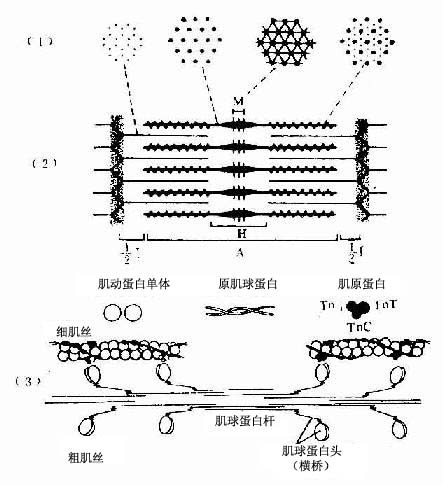
图6-4 骨骼肌肌原纤维超微结构及两种肌丝分子结构模式图(1)肌节不同部位的横切面 ,示粗肌丝与细肌丝的分布(2)一个肌节的纵切面,示两种肌丝的排列(3)粗肌丝与细肌丝的分子结构TnT肌原蛋白T,TnC肌原蛋白C,TnI肌原蛋白I
2.横小管它是肌膜向肌浆内凹陷形成的小管网,由于它的走行方向与肌纤维长轴垂直,故称横小管(transverse tubule,或称T小管)。人与哺乳动物的横小管位于A带与I带交界处,同一水平的横小管在细胞内分支吻合环绕在每条肌原纤维周围(图6-5)。横小管可将肌膜的兴奋迅速传到每个肌节。
3.肌浆网 肌浆网(sarcoplasmic reticulum)是肌纤维内特化的滑面内质网,位于横小管之间,纵行包绕在每条肌原纤维周围,故又称纵小管(图6-5)。位于横小管两侧的肌浆网呈环行的扁囊,称终池(terminal cisternae),终池之间则是相互吻合的纵行小管网。每条横小管与其两侧的终池共同组成骨骼肌三联体(triad)(图6-5)。在横小管的肌膜和终池的肌浆网膜之间形成三联体连接,可将兴奋从肌膜传到肌浆网膜。肌浆网的膜上有丰富的钙泵(一种ATP酶),有调节肌浆中Ca2+浓度的作用。
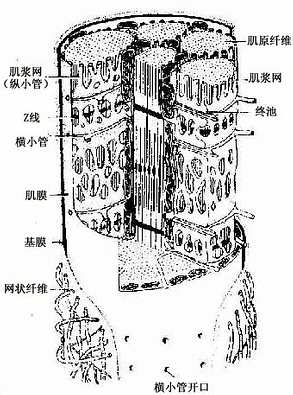
图6-5 骨骼肌纤维超微结构立体模式图
(三)骨骼肌纤维的收缩原理
目前认为,骨骼肌收缩的机制是肌丝滑动原理(sliding filament mechanism)。其过程大致如下:①运动神经末梢将神经冲动传递给肌膜;②肌膜的兴奋经横小管迅速传向终池;③肌浆网膜上的钙泵活动,将大量Ca2+转运到肌浆内;④肌原蛋白TnC与Ca2+结合后,发生构型改变,进而使原肌球蛋白位置也随之变化;⑤原来被掩盖的肌动蛋白位点暴露,迅即与肌球蛋白头接触;⑥肌球蛋白头ATP酶被激活,分解了ATP并释放能量;⑦肌球蛋白的头及杆发生屈曲转动,将肌动蛋白拉向M线(图6-6);⑧细肌丝向A带内滑入,I带变窄,A带长度不变,但H带因细肌丝的插入可消失(图6-7),由于细肌丝在粗肌丝之间向M线滑动,肌节缩短,肌纤维收缩;⑨收缩完毕,肌浆内Ca2+被泵入肌浆网内,肌浆内Ca2+浓度降低,肌原蛋白恢复原来构型,原肌球蛋白恢复原位又掩盖肌动蛋白位点,肌球蛋白头与肌动蛋白脱离接触,肌则处于松驰状态。
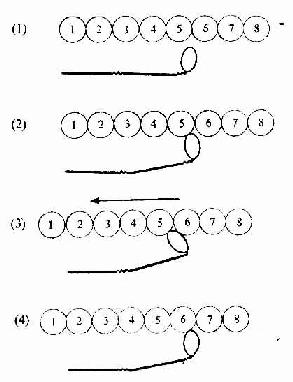
图6-6 骨骼肌纤维收缩的分子结构图解
(1)肌纤维未收缩时,肌球蛋白分子头部未与肌动蛋白接触
(2)肌纤维收缩时,肌球蛋白头部与肌动蛋白位点接触,ATP分解发,释放能量
(3)肌球蛋白头部向M线方向转动,使肌动蛋白丝部向A带滑入
(4)新的接触重新开始

图6-7 骨骼肌纤维收缩时 肌节结构变化图解 (1)肌纤维舒张, (2)肌纤维收缩