神经元(neuron)的形态多种多样,但都可分为胞体(soma)和突起(neurite)两部分(图7-1)。胞体的大小差异很大,小的直径仅5~6μm,大的可达100μm以上,突起的形态、数量和长短也很不相同。神经元突起又分树突(dendrite)和轴突(axon)两种。树突多呈树状分支,它可接受刺激并将冲动传向胞体;轴突呈细索状,末端常有分支,称轴突终末(axon terminal),轴突将冲动从胞体传向终末。通常一个神经元有一个至多个树突,但轴突只有一条(图7-2)。神经元的胞体越大,其轴突越长。
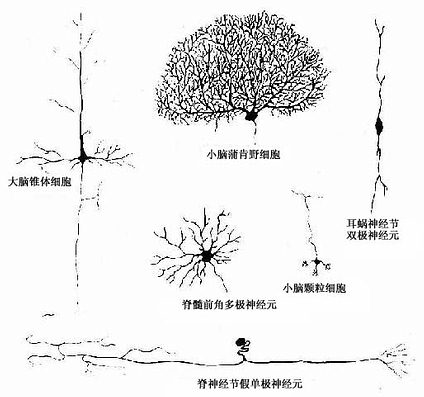
图7-1 神经元的几种主要形态类型
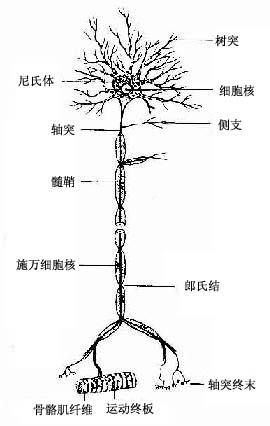
图7-2 运动神经元模式图
(一)神经元的分类
神经元有几种分类法。根据突起的多少可将神经元分为三种:①多极神经元(multipolar neuron),有一个轴突和多个树突;②双极神经元(bipolar neuron),有两个突起,一个是树突,另一个是轴突;③假单极神经元(pseudounipolar neuron),从胞体发出一个突起,距胞体不远又呈“T”形分为两支,一支分布到外周的其他组织的器官,称周围突(peripheral process);另一支进入中枢神经系统,称中枢突(central process)(图7-3)。假单极神经元的这两个分支,按神经冲动的传导方向,中枢突是轴突,周围突是树突;但周围突细而长,与轴突的形态类似,故往往通称轴突。
根据轴突的长短,神经元可分为:①长轴突的大神经元,称GolgiⅠ型神经元,最长的轴突达1m以上;②短轴突的小神经元,称GolgiⅡ型神经元,轴突短的仅数微米。
根据神经元的功能又可分:①感觉神经元(sensory neuron),或称传入神经元(afferent neuron)多为假单极神经元,胞体主要位于脑脊神经节内,其周围突的末梢分布在皮肤和肌肉等处,接受刺激,将刺激传向中枢。②运动神经元(motor neuron),或称传出神经元(efferent neuron)多为多极神经元,胞体主要位于脑、脊髓和植物神经节内,它把神经冲动传给肌肉或腺体,产生效应。③中间神经元(interneuron),介于前两种神经元之间,多为多极神经元(图7-3)。动物越进化,中间神经元越多,人神经系统中的中间神经元约占神经元总数的99%,构成中枢神经系统内的复杂网络。
根据神经元释放的神经递质(neurotransmitter),或神经调质(neuromodulator),还可分为:①胆碱能神经元(cholinergic neuron);②胺能神经元(aminergic neuron);③肽能神经元(peptidergic neuron);④氨基酸能神经元。
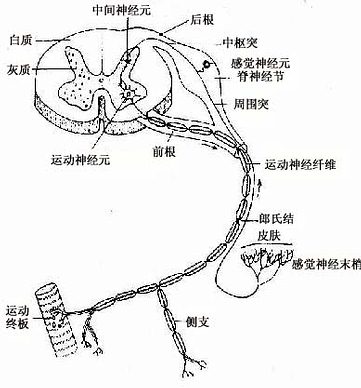
图7-3 脊髓及脊神经,示三种神经元的关系
(二)神经元的结构
1.细胞膜 神经元的细胞膜是可兴奋膜(excitable membrane),它在接受刺激、传播神经冲动和信息处理中起重要作用。通常是神经元的树突膜和胞体膜接受刺激或信息,轴突膜(轴膜)传导神经冲动。神经元细胞膜的性质决定于膜蛋白的种类、数量、结构和功能,其中有些膜蛋白是离子通道(ionic channel)、按所通过的离子分别命名为钠信道、钾信道或钙信道等;还有一些膜蛋白是受体(receptor),可与相应的化学物质(神经递质)结合,使离子通道开放。目前认为,控制离子通道的开闭存在一种闸门机制(gating mechanism),有些通道是受电刺激而开放的,称电位门控通道(voltage-gated channel),有些是当化学物质与受体结合时才开放的,称化学门控通道(chemically-gated channel)。还有一些通道不受上述机制控制,而是经常开放着的。一般是轴膜富含电位门控通道,树突膜和胞体膜主要是化学门控通道。
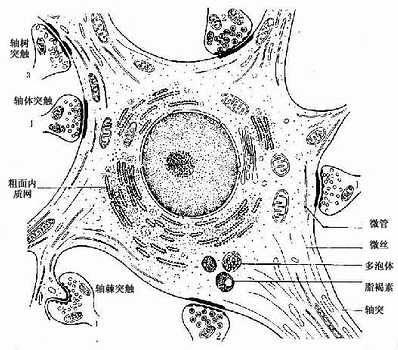
图7-4 多极神经元及其突触超微结构模式图
1.突触扣结内有圆形清亮小泡,内含乙酰胆碱
2、突触扣结内有颗粒型小泡,内含单胺类
3、突触扣结内有扁平清亮小泡,内含甘氨酸等
2.胞体 神经元胞体是细胞的营养中心。胞体的中央有一个大而圆的细胞核,核异染色质少,故着色浅,核仁大而明显。胞体的细胞质称核周质(perikaryon),含有较发达的粗面内质网、游离核糖体、微丝、神经丝和微管以及高尔基复合体等(图7-4)。粗面内质网常呈现规则的平行排列,游离核糖体分布于其间,它们在光镜下呈嗜碱性颗粒或小块,称尼氏体(Nissl bodies)。大神经元尤其是运动神经元的尼氏体丰富而粗大,呈斑块状(图7-5);小神经元的尼氏体则较小而少。大神经元胞体内含大量尼氏体和发达的高尔基复合体,表明细胞具有合成蛋白质的旺盛功能。合成的蛋白质包括复制细胞器所需蛋白质和产生神经递质有关的酶等。神经丝(neurofilament)直径约为10nm,是中间丝的一种,常集合成束,微管直径约25nm,它常与神经丝交叉排列成网,并伸入树突和轴突内,构成神经元的细胞骨架(cytoskeleton),参与物质运输。在银染色切片中,神经丝和微管呈棕黑色细丝,又称神经原纤维(neurofibril)(图7-6)。胞体内还含有色素,最常见的是棕黄色的脂褐素(lipofuscin),并随年龄而增多。

图7-5 神经元胞体光镜结构,示尼氏体
图7-6 神经元银染色,示神经原纤维
某些神经元,如下丘脑具内分泌功能的分泌神经元(secretory neuron),胞体内含直径100~300nm的分泌颗粒,颗粒内含肽类激素(如加压素、催产素等)。
3.树突 树突内的结构与核周质基本相似。在树突分支上常见许多棘状的小突起,称树突棘(dendritic spine)。树突棘是神经元之间形成突触的主要部位,电镜下可见树突棘内有2~3层滑面内质网形成的板层,板层间有少量致密物质,称此为棘器(spine apparatus)(图7-7)。树突棘的数量及分布因不同神经元而异,并可随功能而改变。在大脑皮质锥体细胞和小脑皮质蒲肯野细胞的树突上,树突棘数量最多而明显,一个蒲肯野细胞的树突棘可多达10万个以上。树突的功能主要是接受刺激,树突棘和树突使神经元的接受面大为扩大。
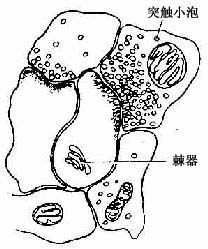
图7-7 树突棘及棘器模式图
4.轴突 轴突通常自胞体发出,但也有从主树突干的基部发出。胞体发出轴突的部位常呈圆锥形,称轴丘(axon hillock),光镜下此区无尼氏体,染色淡(图7-5)。轴突的长短不一,短者仅数微米,长者可达一米以上。轴突一般比树突细,全长直径较均一,有侧支呈直角分出。轴突表面的细胞膜称轴膜(axolemma),内含的胞质称轴质(axoplasm)。轴质内有大量微管和神经丝,此外还有微丝、线粒体、滑面内质网和一些小泡等。微管与神经丝均很长,沿轴突长轴平行排列。微丝较短,主要分布于轴膜下,常与轴膜相连。电镜观察轴突冷冻蚀刻标本,可见微丝、微管和神经丝之间均有横桥连接,构成轴质中的网架结构。轴突内无尼氏体和高尔基复合体,故不能合成蛋白质,轴突成分的更新及神经递质全成所需的蛋白质和酶,是在胞体内合成后输送到轴突及其终末的。
轴突的主要功能是传导神经冲动。神经冲动的传导是在轴膜上进行的,轴突起始段轴膜的电兴奋性阈较胞体或树突低得多,故此处常是神经元发生冲动的起始部位。轴突起始段长约15~25μm,电镜下见轴膜较厚,膜下有电子密度致密层。
轴突内的物质运输称轴突输送(axonal transport)。神经元胞体内新合成的微管、微丝和神经丝组成的网架缓慢地移向轴突终末(0.1~0.4mm/天),称此为慢速输送。另外还有一种快速双向的轴突输送(100~400mm/天)。轴膜更新所需的蛋白质、含神经递质的小泡及合成递质所需的酶等,由胞体输向终末,称快速顺向轴突输送。轴突终末代谢产物或由轴突终末摄取的物质(蛋白质、小分子物质或由邻细胞产生的神经营养因子等)逆行输向胞体,称快速逆向轴突输送(图7-8)。某些微生物或毒素(如破伤风毒素、狂犬病毒)进入轴突终末,也可通过逆行性运输迅速侵犯神经元胞体,新近的研究表明,微管在轴突输送中起重要作用,微管与轴质中的动力蛋白(dynein)或激蛋白(kinesin)相互作用,可推动小泡向一定方向移动。此外微丝也与轴突输送作用有关。
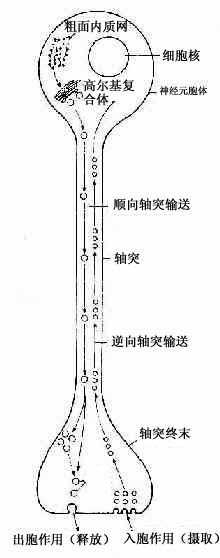
图7-8 双向轴突输送示意图