溶组织内阿米巴(Entamoeba histolytica Schaudinn,1903),即痢疾阿米巴,为侵袭型阿米巴病的病原虫,主要寄生于结肠,引起阿米巴痢疾和各种类型的阿米巴病,为全球分布,多见于热带与亚热带。据统计,在全球超过5亿的阿米巴感染者中,侵袭型的年发病率高达4千万例以上,至今每年死于阿米巴病的人数不少于4万,当前在医学上的重要性已被认为仅次于疟疾与血吸虫病。
人类对阿米巴病早有认识与记载。古希腊希波克拉蒂对痢疾的潜在传染性和并发症-肝泻(hepatic flux)已有明确论述;我国古代医书《内经素问》《伤寒论》等也记有“下痢”,“赤痢”,“疫痢”等有关鉴别诊断的临床经验。阿米巴痢疾的病原体则由俄罗斯医师 лйщ于1975年首先发现,至1903年由德国著名微生物学家Schaudinn定名。对于溶组织内阿米巴的生物学、种群毒力及其致病作用和机制的研究已历经100多年,直到20世纪60年代末美国学者Diamond开拓无共生物纯培养之后,才为细胞及分子水平的生物、生化及免疫学研究提供了高纯高的虫体材料,从而取得新的进展。虽然还留有不少有待深入探讨的问题,对于持续了大半个世纪有关种株与毒力的争论热点,已有初步结论,从生化构成、免疫原性以及基因分析确认了英国学者Brumpt早在1925年即已提出的善于致病与非致病品系属于不同种群的论断。
形态与生活史
近代研究表明,溶组织内阿米巴应是一个包含两种在形态学难于区分而致病力显著不同的种群复合体,即存在着侵袭型和非侵袭型(或称致病和非致病型或弱致病型)两个不同的种。长期以来,人们试图以此解释阿米巴感染的低发病率(<10%)现象,但只有通过70年代末期以来从生化、免疫以及遗传研究的不断探索,终于获得了细胞与分子生物学的确凿证据,并根据研究提供的信息的数据,Diamond和Clark(1993)确认这两个种的存在,重新描述了沿用Entamoeba histolytica Schaudinn,1903学名的侵袭型种的形态学,用以区分非致病种,Entamoeba dispar Brumpt,1925。鉴于这两个种的区分目前仅借助于生化与基因分析手段,对于溶组织内阿米巴的有关生物学描述,仍须借鉴传统的复合群体概念。
溶组织内阿米巴的生活史有滋养体、囊前期、包囊和囊后滋养体各期(图10-1,2),其中囊前期和囊后滋养体分别是滋养体转化为包囊和包圳转化为滋养体的短暂过渡期,仅出现于肠道寄生阶段。整个生活史过程仅须一种哺乳类宿主,人是主要的适宜宿主。猿、猕猴、犬、猪、鼠均有自然或实验感染的报道,但并无重要流行病意义。
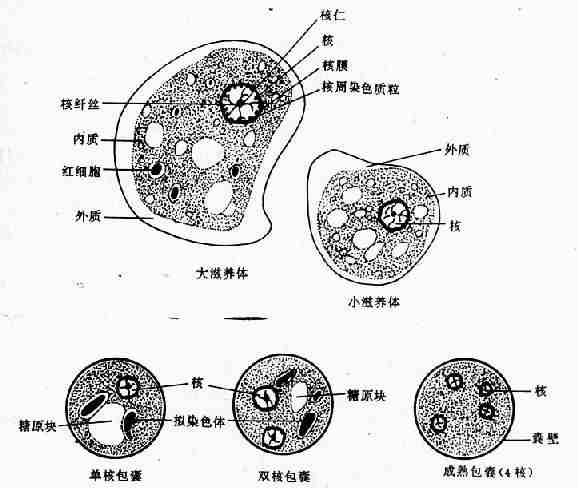
图10-1 溶组织内阿米巴 (铁苏木素染色)
滋养体期系该阿米巴的基本生活型,通常在结肠腔内以二分裂法繁殖。直径10~40µm,个体大小随种系而有差异,可大至60µm以上。在光镜下观察活体,可见较白细胞稍大的折光性活动小体,在适宜温度下运动活泼,常伸出单一伪足作定向阿米巴运动。扫描电镜观察,可见在表膜的一个部位附有许多小泡,为伪尾区,可能是一个排废部位。滋养体需经固定染色后方可辨明内部结构。最常用铁苏木素染色,在高倍放大的光镜下,胞质可辨认较透明的外质和颗粒状内质,内质含一典型泡状核,直径4~7µm,在不着色的纤薄核膜内缘有排列整齐的单层染色质粒(chromatin granules),有一个位于正中或稍偏位的粒状核仁,核仁与核膜之间隐约可见网状核纤丝。典型的核结构有助于虫种鉴别;胞质内含食物泡及吞噬的红细胞。电镜下,滋养体的外被可见由两层电子致密层组成的典型单位膜结构及外层绒毛状糖萼外被。后者的厚度在肠壁病灶分离的滋养体常数倍于无共生物培养的个体。表膜还分布有许多含肌动蛋白的丝状突起为丝状伪足,可证明参与阿米巴的侵袭机制。扫描电镜下的表膜还可见大小不一的杯状吞噬微口(phagocytic stomata),司吞噬与吞饮的功能。内质中可见众多的食物泡、空泡和大小不等的糖原粒,另有微管、溶酶体及小泡状或管状内质网,迄今未发现有线粒体、糙面内质网和典型的高尔基复合体。