(一)DNA超螺旋
双螺旋DNA进一步扭曲盘绕则形成其三级结构,超螺旋是DNA三级结构的主要形式。自从1965年Vinograd等人发现多瘤病毒的环形DNA的超螺旋以来,现已知道绝大多数原核生物都是共价封闭环(covalentlyclosed circle,CCC)分子,这种双螺旋环状分子再度螺旋化成为超螺旋结构(superhelix或supercoil),如图15-11所示。有些单链环形染色体(如φ×174)或双链线形染色体(如噬菌体入),在其生活周期的某一阶段,也必将其染色体变为超螺旋形式。对于真核生物来说,虽然其染色体多为线形分子但其DNA均与蛋白质相结合,两个结合点之间的DNA形成一个突环(loop)结构,类似于CCC分子,同样具有超螺旋形式。超螺旋按其方向分为正超螺旋和负超螺旋两种。真核生物中,DNA与组蛋白八聚体形成核小体结构时,存在着负超螺旋。研究发现,所有的DNA超螺旋都是由DNA拓扑异构酶产生的。
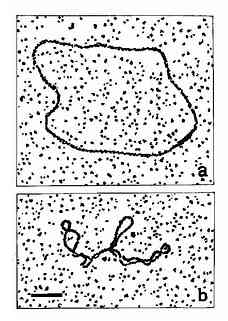
图15-11 Open?circular(a) andsupercoiled (b) forms of PM2 virus DNA,Bar
represents 0.2μm.(By courtesy ofDr Lesley Coggins)
(二)染色质和核小体
1.染色质
真核生物的染色体(chromasome)在细胞生活周期的大部分时间里都是以染色质(chromatin)的形式存在的。染色质是一种纤维状结构,叫做染色质丝,它是由最基本的单位枣核小体(nucleosome)成串排列而成的。DNA是染色体的主要化学成分,也是遗传信息的载体,约占染色体全部成分的27%,另外组蛋白和非组蛋白占66%,RNA占6%。
组蛋白(histones)是一种碱性蛋白质,等电点一般在PH10.0以上,其特点是富含二种碱性氨基酸(赖氨酸和精氨酸),根据这两种氨基酸在蛋白质分子中的相对比例,将组蛋白分为五种类型(表15-6)。
表15-6 五种组蛋白分子的基本参数
| 种类 | 类型 | 碱性氨基酸 | 酸性
氨基酸 | 碱性氨基酸/酸性氨基酸 | 氨基酸
残基数 | 分子量 | 核小体
上位置 | ||
| Lys | Arg | Lys/Arg | |||||||
| H1 | 极度富含Lys | 29% | 1% | 29 | 5% | 5.4 | 215 | 23 000 | 连接* |
| H2A | 11% | 9% | 1.2 | 15% | 1.4 | 129 | 14 500 | 核心 | |
| H2B | ″ | 16% | 6% | 2.7 | 13% | 1.7 | 125 | 13 774 | ″ |
| H3 | 极度富含Lys+富含Arg | 10% | 13% | 0.78 | 13% | 1.8 | 135 | 15 324 | ″ |
| H4 | ″ | 11% | 14% | 0.79 | 10% | 2.5 | 102 | 11 282 | ″ |
*现已证明:H1是与核小体的核心颗粒相当靠近的,认为它位于连接DNA上,只是一种习惯说法。
五种组蛋白在同一生物的不同组织中完全一样,在不同的真核生物中也很相似。组蛋白对染色体中DNA的包装有十分重要的作用。
2.核小体
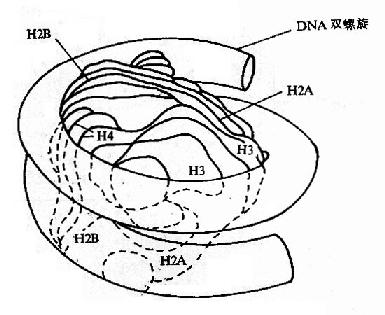
图15-12A DNA的高级结构
核小体是构成染色质的基本结构单位,使得染色质中DNA、RNA和蛋白质组织成为一种致密的结构形式。核小体由核心颗粒(core particle)和连接区DNA(linker DNA)二部分组成,在电镜下可见其成捻珠状,前者包括组蛋白H2A,H2B,H3和H4各两分子构成的致密八聚体(又称核心组蛋白),以及缠绕其上一又四分之三圈长度为146bp的DNA链;后者包括两相邻核心颗粒间约60bp的连接DNA和位于连接区DNA上的组蛋白H1(图15-12),连接区使染色质纤维获得弹性。核小体是DNA紧缩的第一阶段,在此基础上,DNA链进一步折叠成每圈六个核小体,直径30nm的纤维状结构,这种30nm纤维再扭曲成襻,许多襻环绕染色体骨架(Scaffold)形成棒状的染色体,最终压缩将近一万倍。这样,才使每个染色体中几厘米长(如人染色体的DNA分子平均长度为4cm)的DNA分子容纳在直径数微米(如人细胞核的直径为6-7μm)的细胞核中。
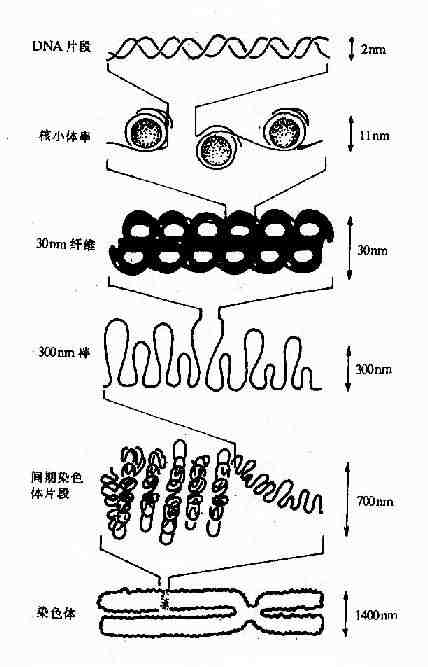
图15-12B DNA的高级结构-从核小体至染色体
核小体的形成以及DNA超螺旋结构与功能的关系还不十分清楚,可能与基因的转录调节控制有关。