DNA做为遗传物质的基本特点就是在细胞分裂前进行准确地自我复制(self?replication),使DNA的量成倍增加,这是细胞分裂的物质基础。1953年Watson和Crick提出DNA双螺旋结构模型指出,DNA是由二条互补的脱氧核苷酸链组成,所以一条DNA链上的核苷酸排列顺序是由图16-1双螺旋DNA的复制另一条决定的。这就说明DNA的复制是由原来存在的分子为模板来合成新的链。曾经有过多种关于DNA复制方式的学说,包括半保留复制,全保留复制以及分散复制等(图16-1)。
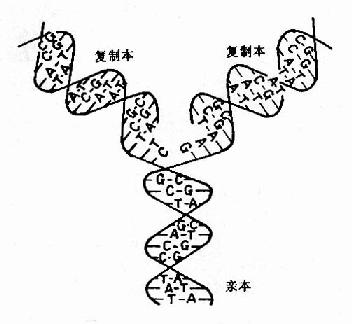
图16-1 双螺旋DNA的复制
一、DNA复制的方式及一般过程:
(一)DNA的半保留复制(semiconservative replication)
Watson和Crick在提出DNA双螺旋结构模型时即推测,DNA在复制时首先两条链之间的氢键断裂两条链分开,然后以每一条链分别做模板各自合成一条新的DNA链,这样新合成的子代DNA分子中一条链来自亲代DNA,另一条链是新合成的,这种复制方式为半保留复制。
1958年Meselson和Stahl利用氮标记技术在大肠杆菌中首次证实了DNA的半保留复制,他们将大肠杆菌放在含有15N标记的NH4Cl培养基中繁殖了15代,使所有的大肠杆菌DNA被15N所标记,可以得到15N桪NA。然后将细菌转移到含有14N标记的NH4Cl培养基中进行培养,在培养不同代数时,收集细菌,裂介细胞,用氯化铯(CsCl)密度梯度离心法观察DNA所处的位置。由于15N桪NA的密度比普通DNA(14N-DNA)的密度大,在氯化铯密度梯度离心(density gradientcentrifugation)时,两种密度不同的DNA分布在不同的区带。
实验结果表明:在全部由15N标记的培养基中得到的15N桪NA显示为一条重密度带位于离心管的管底。当转入14N标记的培养基中繁殖后第一代,得到了一条中密度带,这是15N桪NA和14N-DNA的杂交分子。第二代有中密度带及低密度带两个区带,这表明它们分别为15N14N-DNA和14N14N-DNA。随着以后在14N培养基中培养代数的增加,低密度带增强,而中密度带逐渐减弱,离心结束后,从管底到管口,CsCl溶液密度分布从高到低形成密度梯度,不同重量的DNA分子就停留在与其相当的CsCl密度处,在紫外光下可以看到DNA分子形成的区带。为了证实第一代杂交分子确实是一半15N-DNA-半14N-DNA,将这种杂交分子经加热变性,对于变性前后的DNA分别进行CsCl密度梯度离心,结果变性前的杂交分子为一条中密度带,变性后则分为两条区带,即重密度带(15N-DNA)及低密度带(14N-DNA)。它们的实验只有用半保留复制的理论才能得到圆满的解释(图16-2和16-3)。
 | 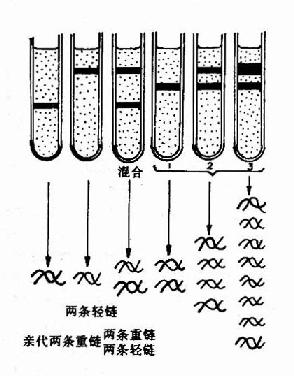 |
| 图16-2 DNA的半保留复制第一代分子含有一条亲代的链(用黑色素示),与另一条新合成的链(用白色表示)配对。在以后的连续复制过程中,原来亲代的两条链仍然保持完整,因此总有两个分子各具有一条原来亲代的链。 | 图16-3 DNA的半保留复制-Meslson?Stahl实验密度梯度离心后的DNA位置:左三管为对照;右三管为实验结果 |
(二)DNA复制的一般过程:
DNA双螺旋是由两条方向相反的单链组成,复制开始时,双链打开,形成一个复制叉(replicative fork,从打开的起点向一个方向形成)或一个复制泡(replicative bubble,从打开的起点向两个方向形成。)两条单链分别做模板。各自合成一条新的DNA链。由于DNA一条链的走向是5′→3′方向,另一条链的走向是3′→5′方向,但生物体内DNA聚合酶只能催化DNA从5′→3′的方向合成。那么,两条方向不同的链怎样才能做模板呢?这个问题由日本学者岗崎先生解决。
原来,在以3′→5′方向的母链为模板时,复制合成出一条5′→3′方向的前导链(leadingstrand),前导链的前进方向与复制叉打开方向是一致的,因此前导链的合成是连续进行的,而另一条母链DNA是5′→3′方向,它作为模板时,复制合成许多条5′→3′方向的短链,叫做随从链(lagging strand),随从链的前进方向是与复制叉的打开方向相反的。随从链只能先以片段的形式合成,这些片段就叫做岗崎片段(Okazaki fragments),原核生物岗崎片段含有1000-2000核苷酸,真核生物一般100?00核苷酸。最后再将多个岗崎片段连接成一条完整的链。由于前导链的合成是连续进行的,而随从链的合成是不连续进行的,所以从总体上看DNA的复制是半不连续复制(图16-4)。
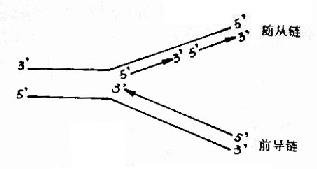
图16-4 DNA的半不连续复制
DNA复制的全部过程可以人为地分成三个阶段,第一个阶段为DNA复制的起始阶段,这个阶段包括起始点,复制方向以及引发体的形成,第二阶段为DNA链的延长,包括前导链及随从链的形成和切除RNA引物后填补空缺及连接岗崎片段。第三阶段为DNA复制的终止阶段。在DNA复制的整个过程中需要30多种酶及蛋白质分子参加,我们将在DNA复制的各个阶段中着重介绍它们的作用。
二、DNA复制的起始阶段:
(一)DNA复制的起始点
很多实验都证明:复制是从DNA分子上的特定部位开始的,这一部位叫做复制起始点(originof replication)常用ori或o表示。细胞中的DNA复制一经开始就会连续复制下去,直至完成细胞中全部基因组DNA的复制。DNA复制从起始点开始直到终点为止,每个这样的DNA单位称为复制子或复制单元(replicon)。在原核细胞中,每个DNA分子只有一个复制起始点,因而只有一个复制子,而在真核生物中,DNA的复制是从许多起始点同时开始的,所以每个DNA分子上有许多个复制子。
DNA复制起始点有结构上的特殊性,例如:大肠杆菌染色体DNA复制起始点Oric由422个核苷酸组成,是一系列对称排列的反向重复序列,即回文结构(palindrome),其中有9个核苷酸或13个核苷酸组成的保守序列,这些部位是大肠杆菌中DnaA蛋白识别的位置,大肠杆菌染色体DNA是环状双链DNA,它的复制是典型的“θ”型复制(由于形状像希腊字母θ)。从一个起点开始,同时向两个方向进行复制,当两个复制方向相遇时,复制就停止。而有些生物的DNA复制起始区是一段富含A·T的区段。这些特殊的结构对于在DNA复制起始过程中参与的酶和许多蛋白质分子的识别和结合都是必须的。
(二)DNA复制的方向:
(1)定点开始双向复制:
这是原核生物和真核生物DNA复制最主要的形式,从一个特定位点解链,沿着两个相反的方向各生长出两条链,形成一个复制泡,用电子显微镜可以观察到复制泡的存在(图16-5)。
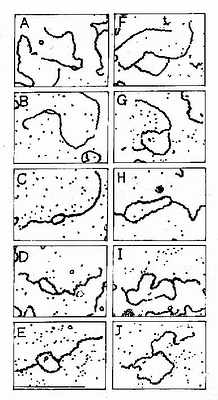
图16-5 SV40DNA;复制泡生长的电镜图谱
(2)定点开始单向复制:
质粒colE1是个典型的例子,复制从一个起始点开始,以同一方向生长出两条链,形成一个复制叉(replication fork)。
(3)两点开始单向复制:
腺病毒DNA的复制是从两个起点开始的,形成两个复制叉,各以一个单一方向复制出一条新链(图16-6)。
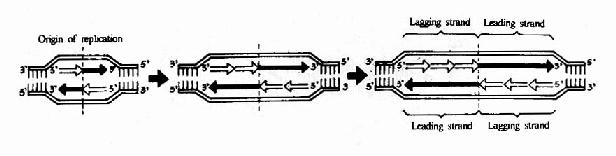
图16-6 DNA的半不连续复制和复制泡的形成
总之DNA复制的起点及方向不仅原核细胞与真核细胞不同,就是同属于原核生物和真核生物的不同种属也有相当大的差异(图16-7)。
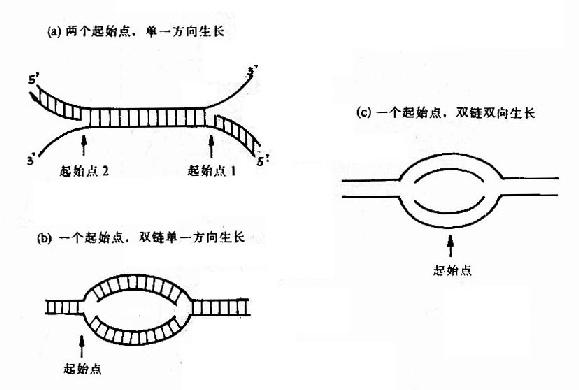
图16-7 DNA链生长方向的三种机制
(三)DNA复制起始引发体的形成及所参与的酶和蛋白质:
1.解链酶(helicase)
DNA开始复制时首先在起始点处解开双链,反应是在一种解链酶(helicase)的催化下进行的。解链酶需要ATP分解供给能量。大肠杆菌中DnaB蛋白就有介链酶活性,与随从链的模板DNA结合,沿5′→3′方向移动,还有一种叫做Rep蛋白和前导链的模板DNA结合沿3′→5′方向移动。解链酶的作用就是打开DNA双链之间的氢键。
2.单链结合蛋白:(single strand binding proteins,SSBP)
它与解开的单链DNA结合,使其稳定不会再度螺旋化并且避免核酸内切酶对单链DNA的水解,保证了单链DNA做为模板时的伸展状态,SSBP可以重复利用(图16-8)。
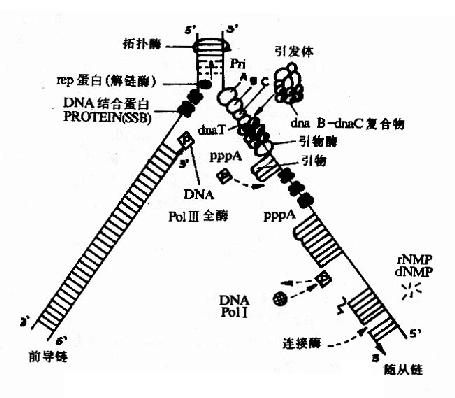
图16-8 大肠杆菌DNA复制叉中复制过程简图
3.引发体的形成:
DNA复制起始的关健步骤是前导链DNA的合成,一旦前导链DNA的聚合作用开始,随从链DNA的合成也随着开始。由于前导链的合成是连续进行的,所以它的起始相对简单,而随从链的合成是不连续进行的,所以引发阶段比较复杂。大肠杆菌的引发前体由Dna B. Dna C和单链结合蛋白组成。
(1)引物酶(primase)
它是一种特殊的RNA聚合酶,可催化短片段RNA的合成。这种短RNA片段一般十几个至数十个核苷酸不等,它们在DNA复制起始处做为引物。RNA引物的3′桹H末端提供了由DNA聚合酶催化形成DNA分子第一个磷酸二酯键的位置。
(2)引发体(primosome)
高度解链的模板DNA与多种蛋白质因子形成的引发前体促进引物酶结合上来,共同形成引发体,引发体主要在DNA随从链上开始,它连续地与引物酶结合并解离,从而在不同部位引导引物酶催化合成RNA引物,在引物RNA的3′桹H末端接下去合成DNA片段,这就是随从链不连续合成的开始。
三、DNA复制的延长阶段以及参与的酶和蛋白质分子:
DNA的复制实际上就是以DNA为模板在DNA聚合酶作用下,将游离的四种脱氧单核苷酸(dATP,dGTP,dCTP,dTTP,简写为dNTP)聚合成DNA的过程。
这是一个非常复杂的酶促反应,需要许多种酶和蛋白质参与,现分别叙述它们在DNA复制中作用。
(一)DNA的聚合反应和DNA聚合酶
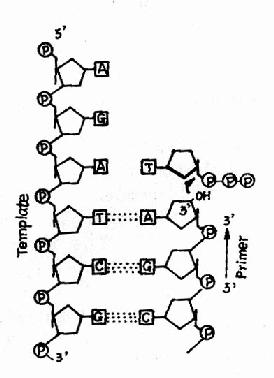
图16-9 DNA聚合酶的作用
1957年,Arthur kornberg首次在大肠杆菌中发现DNA聚合酶Ⅰ,(DNa polymerase Ⅰ,简写DNA polⅠ)后来又相继发现了DNA聚合酶Ⅱ和DNA聚合酶Ⅲ。(DNa polymerase Ⅱ,Ⅲ,简写DNA polⅡ,DNA polⅢ)实验证明大肠杆菌中DNA复制的主要过程靠DNa polⅢ起作用,而DNA polⅠ和DNA polⅡ在DNA错配的校正和修复中起作用。见表16-1。
这种酶的共同性质是:①需要DNA模板,因此这类酶又称为依赖DNA的DNA聚合酶(DNa dependent DNA polymerase, DDDP)。②需要RNA或DNA做为引物(primer),即DNA聚合酶不能从头催化DNA的起始。③催化dNTP加到引物的3′桹H末端,因而DNA合成的方向是5′→3′。图16-9。④三种DNA聚合酶都属于多功能酶,它们在DNA复制和修复过程的不同阶段发挥作用。由于DNA聚合酶Ⅰ是研究得最清楚而且代表了其他DNA聚合酶的基本特点,所以我们着重介绍DNa polⅠ的作用并指出另外二种DNA pol的特殊性:
1.DNA聚合酶Ⅰ:
DNA polⅠ是由一条多肽链组成,分子量为109KD。酶分子中含有一个Zn++,是聚合活性必须的。
大肠杆菌每个细胞中约有400个酶分子,每个酶分子每分钟在37℃下能催化667个核苷酸参入到DNA链中,用枯草杆菌蛋白酶可将此酶水介成两个片段,大片段分子量为76KD,通常称为klenow片段,小片段为34KD。大小片段具有不同的酶活性。
(1)DNA聚合酶的5′→3′聚合活性:
这是DNA聚合酶最主要的活性,按模板DNA上的核苷酸顺序,将互补的dNTP逐个加到引物RNA3′桹H末端,并促进3′桹H与dNTP的5′桺O4形成磷酸二酯键,酶的专一性表现为新进入的dNTP必须与模板DNA碱基配对时才有催化作用,5′→3′聚合活性存在于klenow片段上(图16-9和图16-10)。
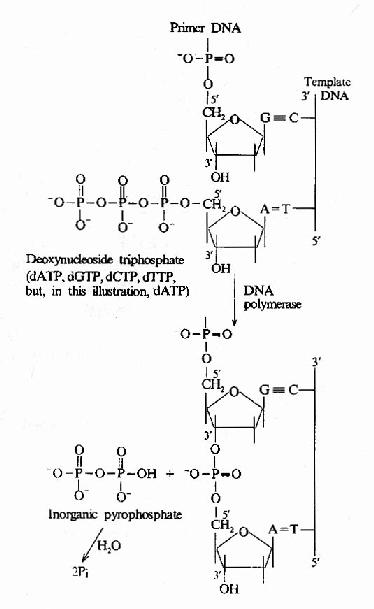
图16-10 DNA聚合酶催化的DNA链延长
(2)DNA聚合酶的3′→5′外切核酸酶活性:
这种酶活性的主要功能是从3′→5′方向识别并切除DNA生长链末端与模板DNA不配对而游离的核苷酸,这种功能称为校对功能,这是保证其聚合作用的正确性不可缺少的,因此对于DNA复制中极高的保真性是至关重要的。
(3)DNA聚合酶的5′→3′外切核酸酶活性:
这种酶活性是从DNA链的5′端向3′末端水解已配对的核苷酸,本质是切断磷酸二酯键,每次能切除10个核苷酸。因此,这种酶活性在DNA损伤的修复中可能起重要作用,对完成的DNA片段去除5′端的RNA引物也是必须的。
DNA polⅠ的5′→3′聚合活性和5′→3′外切酶活性协同作用,可以使DNA一条链上的切口从5′→3′方向移动,这种反应叫做缺刻平移(nick translation),利用此反应可在体外对DNA片段进行放射性磷(α-32PdNTP)的标记制成探针(probe),进行核酸的分子杂交实验,是现代分子生物学的一项重要技术(图16-11)。
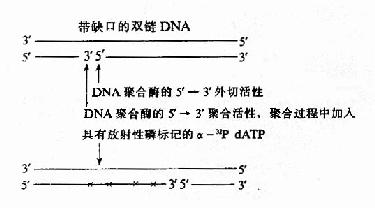
图16-11 缺刻平移标记DNA探针
许多实验证实DNA polⅠ并不是DNA复制过程中的主要酶,它的作用主要与DNA损伤后的修复有关。
2.DNA聚合酶Ⅱ(DNA polⅡ)
此酶分子量为120KD,每个细胞约有100个酶分子,但活性只有DNa polⅠ的5%,它具有5′→3′聚合活性和3′→5′外切活性,而没有5′→3′外切活性,它的作用可能与DNA损伤修复有关。
3.DNA聚合酶Ⅲ(DNA polⅢ)
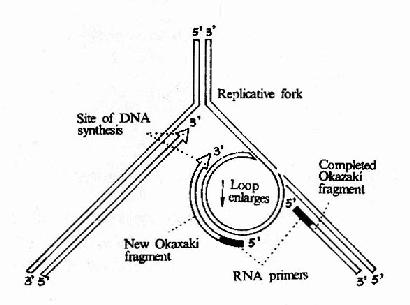
图16-12 DNA聚合酶Ⅲ催化先导链和随从的合成
这是在DNA复制过程中起主要作用的聚合酶,它是由一个多亚基组成的蛋白质分子,其分子量>600kDa整个酶分子形成一个不对称的二聚体,每个大肠杆菌细胞中只有10?0个酶分子,但催化dNTP参入DNA链的速率却是最快的,约为9000核苷酸/每分钟/每个酶分子。这也证明DNa polⅢ是DNA复制过程中主要发挥作用的酶。在大肠杆菌染色体DNA进行复制时,DNA聚合酶Ⅲ全酶并不是单独起作用的,而是与引发体,介链酶等构成一个复制体(replisome)。由于复制体的存在,先导链和随从链可以同时复制。DNa polⅢ是由多亚基组成的不对称二聚体,它可能同时负责先导链和随从链的复制,在φ×174的复制中观察到引发体总是伴随着DNA噜噗(loop)的存在。图16?2可以看到,由于随从链的模板DNA在DNA聚合酶Ⅲ全酶上绕转了180°而形成一个噜噗,因此岗崎片段的合成方向能够与先导链的合成方向以及复制体移动方向保持一致。
随着DNA polⅢ向前移动,先导链的合成逐渐延长的同时,岗崎片段也在不断延长,这一噜噗也在不断扩大。当岗崎片段合成到前一个片段的5′端时,这一大噜噗就释放出来,由于复制叉向前移动又可将另一部分随从链的模板置换出来,由引发体合成新的引物,然后再形成一个小的噜噗,进行新的岗崎片段的合成。由此模型不难看出:随从链的合成需要周期性的引发,因此其合成进度总是与前导链相差一个岗崎片段的长度。岗崎片段完成后,其5′端的RNA引物由DNa polⅠ的5′→3′外切酶活性切除,由此造成的空隙再由DNA polⅠ的5′→3′聚合活性催化dNTP得到填补。所以DNA的复制是在DNa polⅢ和DNApolⅠ互相配合下完成的。
下面列表说明三种大肠杆菌DNA聚合酶的特性
表16-1 大肠杆菌DNA聚合酶特征
| DNA聚合酶Ⅰ | DNA聚合酶Ⅱ | DNA聚合酶Ⅲ | |
| 分子量 | 109KD | 120KD | >600KD |
| 每个细胞中的分子数 | 400 | 17-100 | 10-20 |
| 5′→3′聚合活性 | + | + | + |
| 37℃转化率核苷酸数/酶分子·分钟 | 600 | 30 | 30,000 |
| 5′→3′外切活性 | + | - | - |
| 5′→3′外切活性 | + | + | + |
| 切刻平移活性 | + | - | - |
| 对dNTP亲和力 | 低 | 低 | 高 |
| 功能 | 修复 | 不详 | 复制 |
| 去除引物 | |||
| 填补空缺 |
真核生物DNA聚合酶
真核生物DNA聚合酶有α、β、γ、δ及ε。它们的基本特性相似于大肠杆菌DNA聚合酶,其主要活性是催化dNTP的5′→3′聚合活性,基本特征见表16-2。
表16-2 真核生物DNA聚合酶
| α | β | γ | δ | ε | |
| 亚基数 | 4 | 4 | 4 | 2 | 5 |
| 分子量(KD) | >250 | 36-38 | 160-300 | 170 | 256 |
| 细胞内定位 | 核 | 核 | 线粒体 | 核 | 核 |
| 5′→3′聚合活性 | + | + | + | + | + |
| 3′→5′外切活性 | - | - | - | - | - |
| 功能 | 复制、引发 | 修复 | 复制 | 复制 | 复制 |
真核细胞在DNA复制中起主要作用的是DNA polα,主要负责染色体DNA的复制。DNa polβ的模板特异性是具有缺口的DNA分子,被认为它与DNA修复有关。DNa polγ在线粒体DNA的复制中起作用。DNA polδ不但有5′→3′聚合活性,而且还具有3′→5′外切酶活性,据认为真核生物DNA复制是在DNa polα和DNA polδ协同作用下进行的,前导链的合成靠DNA polδ催化,并且还需要一种细胞周期调节因子椩鲋诚赴?丝乖?proliferatingcell nucleus antigen, PCNA)参与。而随从链的合成靠DNA polα和引发酶配合作用完成。
(二)与超螺旋松驰有关的酶:
DNA复制从起始点开始向一个方向复制时,局部的DNA双链必须打开,主要靠解链酶的作用,打开后的单链还需要单链结合蛋白与其结合,在复制叉向前移动时造成其前方DNA分子所产生的正超螺旋,必须由拓扑异构酶来解决。下面分别介绍它们的作用。
拓扑异构酶(topoisomerase)是一类改变DNA拓扑性质的酶。在体外可催化DNA的各种拓扑异构化反应,而在生物体内它们可能参与了DNA的复制与转录。在DNA复制时,复制叉行进的前方DNA分子部分产生有正超螺旋,拓扑酶可松驰超螺旋,有利于复制叉的前进及DNA的合成。DNA复制完成后,拓扑酶又可将DNA分子引入超螺旋,使DNA缠绕、折叠,压缩以形成染色质。DNA拓扑异构酶有Ⅰ型和Ⅱ型,它们广泛存在于原核生物及真核生物中。表16-3
表16-3 大肠杆菌和真核生物中的拓扑异构酶
| 类型 | 作用 | 对超螺旋的作用 |
| Ⅰ型拓扑异构酶 | ||
| 大肠杆菌 | 切开一股DNA链 | 松驰负超螺旋 |
| 真核生物 | 切开一股DNA链 | 松驰正,负超螺旋 |
| Ⅱ型拓扑异构酶 | ||
| 大肠杆菌 | 切开二股DNA链 | 构驰正超螺旋; |
| 依赖ATP | 引入负超螺旋, | |
| 解环连等 | ||
| 真核生物 | 切开二股DNA链 | 松驰正超旋, |
| 依赖ATP | 但不能引入负超螺旋 |
拓扑异构酶Ⅰ(TopoⅠ)的主要作用是将环状双链DNA的一条链切开一个口,切口处链的末端绕螺旋轴按照松驰超螺旋的方向转动,然后再将切口封起来。这就使DNA复制叉移动时所引起的前方DNA正超螺旋得到缓解,利于DNA复制叉继续向前打开。拓扑异构酶Ⅰ除上述作用外,对环状单链DNA还有打结或解结作用,对环状双链DNA的环连或解连以及使环状单链DNA形成环状双链DNA都有作用(图16-13)。
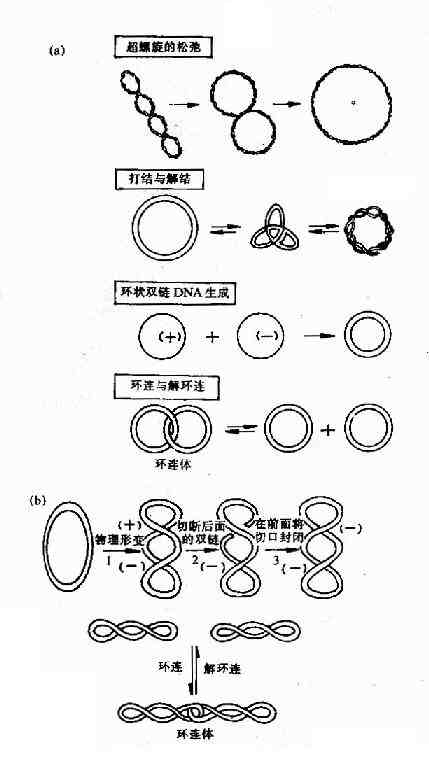
图16-13 拓扑酶Ⅰ及Ⅱ的作用特点
(a)大肠杆菌拓扑酶Ⅰ催化的4种拓扑异构化作用 (b)拓扑酶Ⅱ的作用
拓扑异构酶Ⅱ(TopoⅡ)是在大肠杆菌中发现的,曾被称为旋转酶(gyrase),它们作用特点是切开环状双链DNA的两条链,分子中的部分经切口穿过而旋转,然后封闭切口,TopoⅡ还可使DNA分子从超螺旋状态转变为松驰状态,此反应不需要ATP参与。DNA复制完成后,TopoⅡ在ATP参与下,DNA分子从松驰状态转变为负超螺旋。此外,TopoⅡ催化的拓扑异构化反应还有环连或解环连,以及打结或解结。
四、DNA复制的终止阶段
DNA在复制过程中,合成出的前导链为一条连续的长链。随从链则是由合成出许多相邻的片段,在连接酶的催化下,连接成为一条长链。连接作用是在连接酶催化下进行的。
连接酶(ligase)的作用是催化相邻的DNA片段以3′、5′-磷酸二酯键相连接。连接反应中的能量来自ATP(或NAD+)。连接酶先与ATP作用,以共价键相连生成E桝MP中间体。中间体即与一个DNA片段的5′-磷酸相连接形成E-AMP-5′-DNA。然后再与另一个DNA片段的3′-OHH末端作用,E和AMP脱下,两个DNA片段以3′、5′磷酸二酯键相连接。随从链的各个DNA片段就是这样连接成一条DNA长链(图16-14)。?
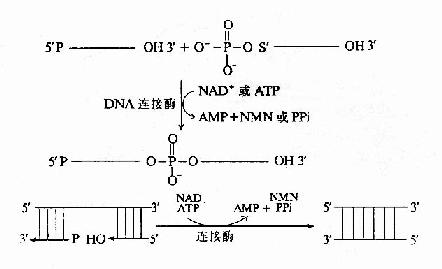 | 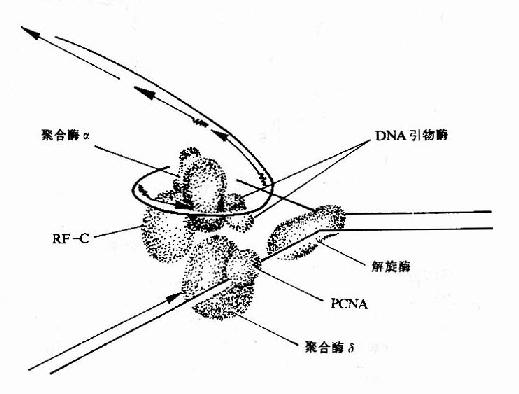 |
| 图16-14 连接酶的催化反应 | 图16-15 真核生物DNA复制叉结构示意图 |
已有研究证明大肠杆菌染色体DNA具有复制终止位点,此处可以结合一种特异的蛋白质分子叫做Tus,这个蛋白质可能是通过阻止解链酶(Helicase)的解链活性而终止复制的。详细的机制还不完全清楚。
DNA复制完成后,靠拓扑酶将DNA分子引入超螺旋结构。
五、真核生物DNA复制的特点:
DNA复制的研究最初是在原核生物中进行的,有些原核生物的DNA复制已经搞得很清楚。真核生物比原核生物复杂得多,但DNA复制的基本过程还是相似的。在这里我们主要讨论一些重要的区别。
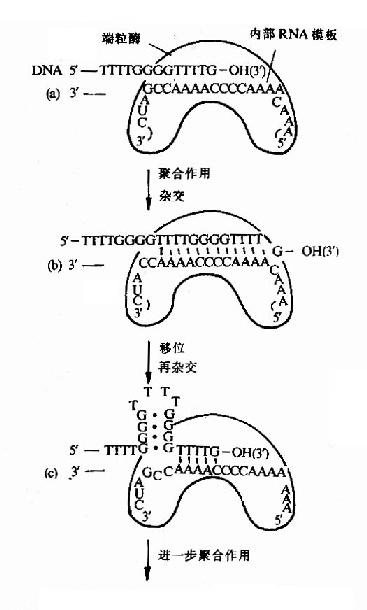
图16-16 端粒酶催化端区TG链的合成
1.与原核生物不同,真核生物DNA复制有许多起始点,例如酵母S.cerevisiae的17号染色体约有400个起始点,因此,虽然真核生物DNA复制的速度(60核苷酸/每秒钟)比原核生物DNA复制的速度(E.coli1700核苷酸/每秒钟)慢得多,但复制完全部基因组DNA也只要几分钟的时间。
2.SV40病毒DNA主要依靠宿主细胞中的DNA复制体系进行DNA的复制,这是了解真核生物DNA复制的体外模型。在真核生物DNA复制叉处,需要两种不同的酶。DNA聚合酶α(polα)和DNA聚合酶δ(polδ)。polα和引物酶紧密结合,在DNA模板上先合成RNA引物,再由polα延长DNA链,这种活性还要复制因子C参与。同时结合在引物模板上的PCNA(增殖细胞核抗原Proliferating cell nuclear antigen)此时释放了polα,然后由polδ结合到生长链3′末端,并与PCNA结合,继续合成前导链。而随从链的合成靠polα紧密与引物酶结合并在复制因子C帮助下,合成岗崎片段(图16-15)。
3.由于真核生物染色体是线性DNA,它的两端叫做端区(telomeres),端区是由重复的寡核苷酸序列构成的。例如酵母的端区重复序列是5′G(1?)T(3)3′。前面讲到所有生物DNA聚合酶都只能催化DNA从5′→3′的方向合成,因此当复制叉到达线性染色体末端时,前导链可以连续合成到头,而由于随从链是以一种不连续的形式合成岗崎片段,所以不能完成线性染色体末端的复制,如果这个问题不解决,真核生物在细胞分裂时DNA复制将产生5′末端隐缩,使DNA缩短,近十多年的研究表明,真核生物体内都存在一种特殊的反转录酶叫做端粒酶(telomerase),它是由蛋白质和RNA两部分组成的,它以自身的RNA为模板,在随从链模板DNA的3′桹H末端延长DNA,再以这种延长的DNA为模板,继续合成随从链(图16-16)。
由此可见端粒酶在保证染色体复制的完整性上有重要意义。