细胞癌基因的激活是指原本不致癌c-onc在特定的情况下转变成致癌性的,大体上有以下几种激活方式。
1、插入激活 例如逆转录病毒MoSV感染鼠类成纤维细胞后,病毒基因组的LTR整合到细胞癌基因c-mos邻近处,使c-mos处于LTR的强启动子和增强子作用之下而被激活,导致成纤维细胞转化为肉瘤细胞,又如禽类白细胞增生病毒ALV的E成分整合到鸡细胞基因组c-myc附近。可使c-myc激活。因此在基因治疗中使用逆转录病毒载体时必需考虑细胞癌基因的插入激活问题。
2、突变激活 典型的是各种ras基因的激活,参看表22-2。
表22-2 正常Ras蛋白和转化Ras蛋白中氨基酸的变异
| ras基因 | 氨基酸的位置 | |||
| 12 | 13 | 59 | 61 | |
| H-ras-1来源的
正常人c-H-ras-1 人EJ膀胱癌症 人HS242乳癌 鼠Harvey病毒v-H-ras | Gly
GGC Val GTC Arg CGC | Gly | Ala
Thr | Gln
CAG Leu CTG |
| K-ras-2来源的
正常人c-K-ras-2 人Caalu肺癌 人SW480结肠癌 鼠Kirsten病毒v-K-ras | Gly
Lys TGT Val GTT Ser | Gly | Ala
Thr | Gln |
| N-ras来源的
正常人N-ras 人神经母细胞瘤 人早幼粒细胞白血病细胞系 人纤维肉瘤 人AML33 | Gly | Gly
Asp | Ala | Gln
Lys Lys Lys |
ras基因的表达产物Ras是一种小分子G蛋白,在信号转导中起重要作用,正常Ras的作用因其自身的GTP酶活性而受到严格控制,而突变了的Rad其GTP酶活性下降或丧失,失去了原有控制,致使增殖信号持续作用,细胞发生恶性转化,如图22-3所示。
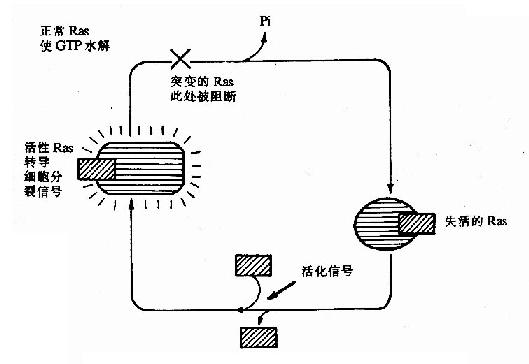
图22-3 Ras与GTP/GDP的相互作用及突变的RasR的作用
3、基因扩增 已发现人类肿瘤细胞中扩增的细胞癌基因如下表。
表22-3 人类肿瘤细胞中扩增的细胞癌基因
| c-onc | 肿瘤 | 扩增倍数 | DM/HSR* |
| c-myc | 早幼粒白血病细胞系HL60 | 20× | + |
| 小细胞肺癌细胞系 | 5-30× | ? | |
| N-myc | 原发神经母细胞瘤Ⅲ-Ⅳ级及神经母细胞瘤细胞系 | 5-1000× | + |
| 视网膜母细胞瘤 | 10-200× | + | |
| 小细胞肺癌 | 50× | + | |
| L-myc | 小细胞肺癌 | 10-20× | ? |
| c-myb | 急粒AML | 5-10× | ? |
| 结肠癌细胞系 | 10× | ? | |
| c-erbB | 类表皮癌细胞系,原发胶质瘤 | 30× | ? |
| c-K-ras | 原发肺癌,结肠癌,膀胱癌,直肠癌 | 4-20× | ? |
| N-ras | 乳癌细胞系 | 5-10× | ? |
*DM:双微体;HSR:均匀染色区
4、基因重排/染色体易位
典型的如伯基特淋巴瘤细胞的染色体易位t(8:14),致使c-myc激活,参看表22-4和图22-4。
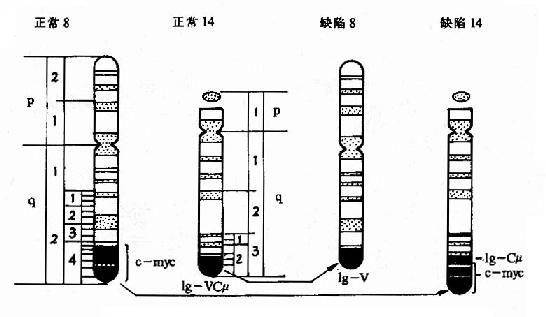
图22-4 Burkitt淋巴瘤常见的染色体易位t(8:14)
表22-4 染色体异常与癌基因重排
| 癌基因 | 染色体定位 | 异常 | 人类肿瘤 |
| c-myc | 8q24 | t(8:14),t(8:22) | Burkitt淋巴瘤 |
| t(2:8) | |||
| bcl-1 | 11q13 | t(11:14) | B细胞淋巴瘤 |
| bcl-2 | 18q21 | t(14:18) | |
| tcl-2 | 11q13 | t(11:14) | T细胞淋巴瘤 |
| c-abI | 9q34 | t(9:22) | 慢粒CML |
| bcr | 22q11 | ph | |
| c-mos | 8q22 | t(8:21) | 急粒AML |
| c-myb | 6q22-24 | t(6:14) | 卵巢癌 |
| c-sis | 22q12 | t(11:22) | Erwing网瘤 |
| blym | 1q32-ter | 缺失,HSR | 神经纤维瘤 |
| c-K-ras | 6q21 | 断裂 | ANLL |
| 6q三体性 | 视网膜母细胞癌 | ||
| c-erbA | 17q21 | 断裂 | ANLL |
已知B淋巴细胞中免疫球蛋白重链基因表达十分活跃,其启动子为强启动子,且在CH-VH之间还有增强子区,c-myc易位后与IG重链基因的调控区为邻,因而被激活。正常情况下,位于c-myc5’端的两个启动子受到c-myc产物的反馈抑制,由此重排时5’端序列有丢失,结果摆脱了抑制而表达增强。
不同的癌基因有不同的激活方式,一种癌基因也可有几种激活方式。例如c-myc的激活就有基因扩增和基因重排两种方式,很少见c-myc的突变;而ras的激活方式则主要是突变,1985年Slamon检测了20种54例人类肿瘤中的15种癌基因,发现所有肿瘤都不止一种癌基因发生改变。细胞转化实验证明,各种癌基因之间存在协同作用。例如,单独v-myc或EJ-ras都不能使大鼠胚胎成纤维细胞转化,但是若将二者共转染PEF,8天后80%的细胞发生变化,那么为什么单独EJ-ras又可使Rat-1细胞转化呢?原因是该细胞并非正常,而是已经永生化了的细胞,如果先用化学诱癌物或射线使正常大鼠原代成纤维细胞永生化,然后再用EJ-ras转染,则可使之转化,因此Weingerg按转染细胞表型的变化将癌基因分为两个类,一类是核内作用的能使细胞永生化的癌基因,例如myc,fos等,另一类是引起细胞恶性表型变化的定位于质膜和胞浆的癌基因,例如ras、erbB、src等。事实表明肿瘤的发生是多步骤,多因素的,不同的癌基因作用于肿瘤发生的不同阶段。
不仅癌基因之间有协同作用,癌基因与抑癌基因之间也存在协同作用。