编码T细胞受体(T cell receptor,TCR)α、β、γ和δ链的基因定位于不同的染色体(表3-6)。人和小鼠δ基因都位于α基因的复合体中,均位于14号染色体;人TCRβ和γ链基因分别位于第7对染色体的长臂和短臂,小鼠β和γ链基因则分别位于第6和13号染色体。
表3-6 TCR多肽链基因定位
| TCR基因 | 染色体定位 | |
| 人 | 小鼠 | |
| α 链 | 14q11.12 | 14 |
| β 链 | 7q32 | 6 |
| γ 链 | 7p15 | 13 |
| δ 链 | 14q11.2 | 14 |
一、TCR基因的结构
TCR基因的结构和重排与Ig基因有许多相似之处。在胚系中,编码TCR多肽链的DNA是由几个分隔开的DNA片段组成,在胸腺细胞中重排后,形成编码一条完整多肽的基因。TCR多肽链可变区基因是由2~4个基因片段通过重排连接在一起。V基因片段编码信号序列和可变区氨基端95~100个氨基酸残基;J基因片段编码可变区羧基端13~23个氨基酸残基。TCRβ、δ链V区基因除V、J基因片段外,还有1~2个D基因片段,编码V与J之间数个氨基酸残基。TCR不同多肽链可变区基因的重排可有V-J、V-D-J或V-D-D-J等几种方式。TCR多肽链C基因片段通常由3~4个外显子所组成,位于J基因片段的下游。与IgC基因片段不同,TCrC基因片段是由数个外显子编码一个结构域,如β链的连接肽(connectingpeptide)是由3个分隔的外显子所编码。
(一)TCRα链基因
TCRα链基因与TCRβ链和Ig基因结构有很大差别。小鼠α链V基因约100个,至少可分为12个家族。人Vα大约在100个左右,长约数百个kb。TCRα基因没有Dα基因片段。人和小鼠α链基因都含有J基因片段(Jα),小鼠约有60个,相互间隔至少500bp。VαJα连接具有多样性。TCRα链基因只有1个Cα基因片段,含有4个外显子。在TCRα链基因座中有TCRδ链基因。TCRα链基因重组信号中长的间隔序列(23bp)3′端靠近Vα,短的间隔序列(12bp)的5′端靠近Jα,这种重组信号的排列方式类似Ig轻链V区的基因,允许Vα基因直接与Jα基因重排,而不需要VD基因片段。
(二)TCRβ链基因
小鼠胚系中未重排的β链基因结构已经搞清,长约数百kb。Vβ基因数目约30个,大多数Vβ基因片段位于Cβ1的上游,至少有1个Vβ(Vβ14)在Cβ2基因3′下游10kb处,与转录的方向相反。人Vβ基因片段约有100个。小鼠和人都有2个Dβ基因片段,12~14bp长,分别位于Jβ1和Jβ2基因片段组上游约500~600bp处。小鼠和人都有2组(cluster)Jβ基因片段,称为Jβ1和Jβ2,与相应的Cβ1和Cβ2基因片段间隔2~3kb。小鼠Jβ1和Jβ2各含7个Jβ基因片段,其中6个有功能,Jβ基因片段相互间隔36~421bp。人Jβ1和Jβ2各含7个有功能的Jβ基因片段。人和鼠TCRβ基因都有2个基本相同的Cβ基因片段,各含4个外显子。Cβ1和Cβ2高度同源,2个Cβ基因片段的编码产物中只有4个氨基酸残基(小鼠)或5个氨基酸(人)不同。
TCRα和β链V基因编码区域包括了相当于Ig的CDR1和CDR2,主要是识别MHC分子和抗原的复合物。CDR3主要是由于V-D-J或V-D-D-J连接的多样性(junctionaldiversity)和广泛分布的N核苷酸插入(Nnucleotide insertion)所造成,其有高度的多样性,可识别众多种类的抗原。
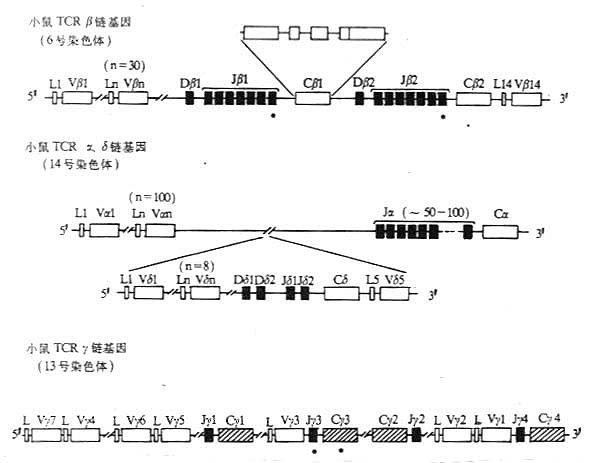
图3-15 小鼠TCR基因的结构
(三)TCRγ链基因
Vγ基因片段数较少,在Balb/c小鼠Vγ基因片段有7个,分为5个家族,其中4个家族各只含1个Vγ,另一家族有3个Vγ。人Vγ有14个,其中8个是有功能的。人和小鼠TCRγ链基因均不含D基因片段。在小鼠,有4个Jγ基因片段,基中Jγ1、Jγ3和Jγ4分别位于3个Cγ基因片段的上游,Jγ3为假基因。在人类共有5个Jγ基因片段,可分为Jγ1和Jγ2两组,其中Jγ1的3个片段位于Cγ1上游,Jγ2的2个片段位于Cγ2基因片段的上游。不鼠有4个Cγ基因片段,其中Cγ3是假基因。人有2个Cγ基因片段,相距约10kb,其中1个Cγ有3个外显子,另一个Cγ含有4个或5个外显子。编码γ链连接肽部分的两个Cγ基因外显子组成存在着差异,有一个Cγ基因片段不编码半胱氨酸,因此造成了TCRγ链在分子量、N-连接糖基化的形式以及是否与TCRδ链之间形成二硫键有所不同。
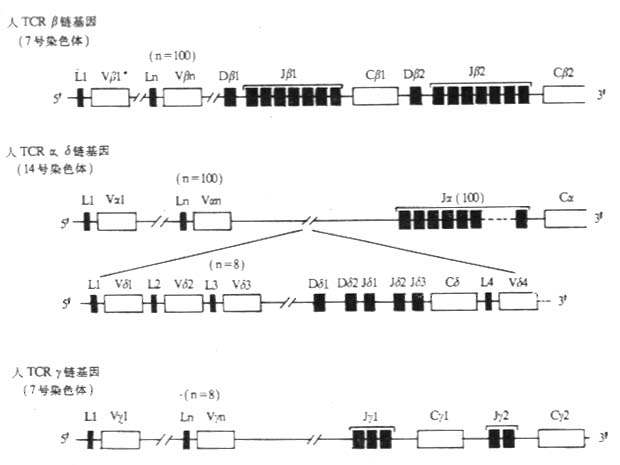
图3-16 人TCR基因的结构
(四)TCRδ链基因
人和小鼠TCr δ链基因结构十分相似,均位于α链基因座内。在Balb/c小鼠,Vδ基因片段数约为8个。在小鼠δ链基因中有2个Dδ、2个Jδ和1个Cδ基因。在胸腺细胞分化过程中,δ链先于α链表达。Vα与Jα重排可导致δ基因复合物中D、J和C基因片段的缺失。在人δ链基因中有8个Vδ基因片段,2个Dδ、3个Jδ和1个Cδ。某些Vδ基因片段是与TCRα链Vα基因共用的。
TCRδ链基因连接信号的排列方式是:每个Vδ基因片段的3′端紧靠着“七聚体-长间隔序列-九聚体”(7-23-9)结构;Dδ基因片段5′端是“九聚体-短间隔序列-七聚体(9-12-7)结构,3′端是“7-23-9”结构;2个Jδ基因片段的5′端是“9-12-7”结构。这种TCRδ链基因重组信号排列的方式使得V、D、J链接可有以下几种不同方式:(1)V-D-J连接,在小鼠胚胎TCRγδ胸腺细胞最常见到Vδ-Dδ2-Jδ的连接,人胚胎胸腺细胞最常见到Vδ-Dδ3-Jδ;(2)V-J的直接连接;(3)大多数成年小鼠TCRγδ胸腺细胞以Vδ-Dδ1-Dδ2-Jδ连接,人妊娠后期胎儿TCRγδ胸腺细胞也存在着Vδ-Dδ1-Dδ2-Jδ的排列方式。Vδ基因位于Vα基因之中,同一个V基因片段可用作Vδ或Vα,如果V基因片段是用作Vδ,多发生V-D(-D)-J重排。
小鼠TCRγδ T细胞γ和δ链配对的,Vδ1几乎总是和Vγ5或Vγ6配对。小鼠TCRγδ阳性树突状表皮细胞(DEC)几乎都是Vγ5与Vδ1配对。在人Vδ2几乎总是与Vγ9配对。
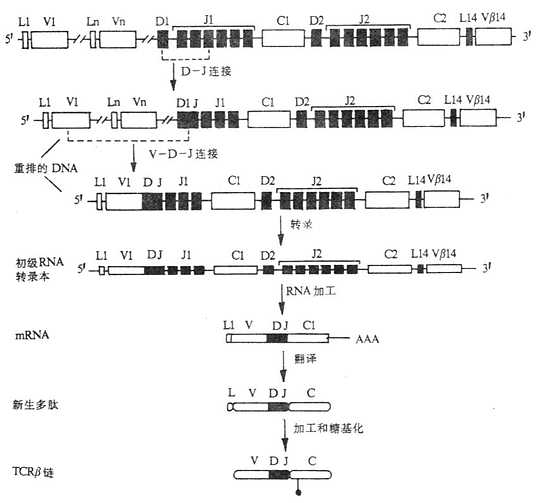
图3-17 小鼠TCRβ基因重排顺序、转录和合成
二、TCR基因的重排
T细胞在胸腺中发育成熟过程中,TCR基因按照一定的顺序发生重排。TCR基因的重排顺序和表达与免疫球蛋白基因的重排和表达十分相似。在基因组中的识别序列包括了一个保守的七聚体和九聚体,七聚体与九聚体之间含有一个不保守的12碱基对或23碱基对间隔序列(spacer sequence)。
TCRβ链基因座的重排要先于α链。首先是一个Dβ片段与一个Jβ片段连接,然后D-J与一个Vβ片段相连,完成了VDJ基因的重组。TCRβ链初级的RNA转录本在重组的VDJ和C基因之间含有内含子。VDJ与Cβ1或Cβ2的重排是随机的,目前尚未发现Cβ类似Ig重链类别转换那样从一个Cβ基因转换到另一个Cβ基因上。在小鼠Cβ2基因的3′端推测有一个增强子。
TCRβ链基因的功能性重排和表达,诱导了TCRα基因的重排。α链基因的重排与β链相似,由于α链基因不存在D片段,故α链基因只有V-J的连接。小鼠Vα转录起始点的5′端有一个启动子,Cα基因的3′端也有一个增强子。此外,靠近α链增强子处有“静息顺序”(silencersequences),这些顺序可抑制非T细胞或者是TCRγδT细胞中α链的转录。由于α链只有一个Cα基因,因此一旦初始的转录本形成后,经过RNA加工只能产生一种完整的α链mRNA。
如同Ig基因的重排和表达,TCRα和β链基因的重排和表达也有等位基因排斥现象(al-lelicexclusion),如果在一条染色体上TCR基因的重排是有效的,那末就可以抑制另一条染色体相应等位基因座的重排。当一条染色体上α链或β链基因座的重排、转录或翻译无效时,则另一条染色体上相应α或β链相应的等位基因座开始发生重排。如果两条染色体上TCRα或β链基因重排都无效,则未成熟的T细胞死亡。
三、T细胞库的多样性
推测T细胞库(T cell repertoire)中多样性在1010以上,其多样性的原因与Ig相似,主要有以下几方面。
1.胚系中有众多的V、D和J基因片段 尽管TCRα和β基因中V基因片段数比Ig基因少,但J基因片段明显比Ig基因为多,如小鼠中约50~100个Jα和至少12个Jβ基因片段,而IgH和κ链基因座中J基因片段各只有4个。
2.连接处连接的多样性 与IgVDJ连接的多样性相似。
3.N区的插入 N区插入在Ig基因重排中只发生在重链基因,而TCRα和β链基因均可发生N区的插入。此外,在TCRα和β链基因中都发现一种称之为“易变性”(flexibility)的现象,如有一个以上Vα基因3′端可以连接到Jα片段。
4.TCRα与β链的随机配对 人TCRαβ库多样性可推算为:α链(Vα(100)×Jα(100))×β链(Vβ(100)×Dβ(2)×Jβ(14))=2.8×107,这种多样性尚未包括N区插入所扩展的多样性。
(金伯泉)