一、HLA的基因组成
人类的MHC称为HLA复合体,位于第6对染色体的短臂上,长度为4分摩(centimorgan,cM),约为4000kb。整个复合体上有近60个基因座,已正式命名的等位基因278个。根据编码分子的特性不同,可将整个复合体的基因分成三类:Ⅰ类、Ⅱ类和Ⅲ类基因(图6-2)。
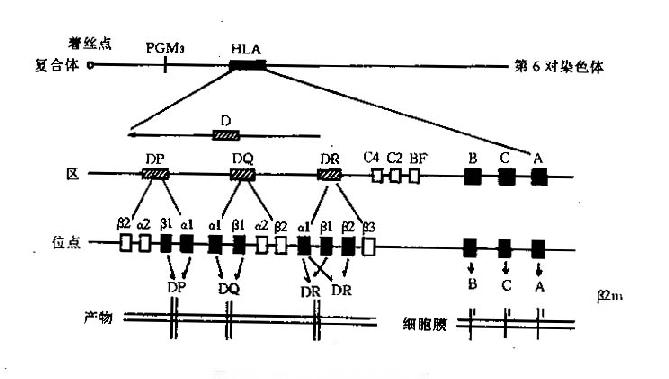
图6-2HLA基因结构示意图
1.类基因区位于着丝点的远端,主要包括HLA-A、B、C三个位点;新近又提出E、F、G、H、K和L位点。
2.类基因区位于着丝点的近端,是结构最为复杂的一个区,主要由DR、DQ、DP三个亚区构成,每个亚区又有若干个位点。新近又鉴定了DO、DZ、DX三个亚区。
3.类基因区含有编码补体成分C2、C4、B因子及TNF、热休克蛋白和21羟化酶的基因。
4.非HLA基因这些基因位于HLA区域内,其功能与HLA相关;目前已经命名的有两类:LMP(largemultifunctionalprotease,或lowmolicularweightpolypeptides)和TAP(transporterassociatedwithantigenprocessing,或transporterofantigenpeptides)。LMP为蛋白酶体相关基因,由LMP2和LMP7组成;TAP为ABC转运蛋白基因,包括TAP1和TAP2;它们的功能可能与抗原的处理和递呈有关。
二、HLA的多态性
HLA复合体是人体最复杂的基因系统,呈高度的多态性,主要原因之一是由于HLA复合体的复等位基因所致。
遗传学上将某一个体同源染色体上对应位置的一对基因称为等位基因(alleles);当群体中位于同一位点的等位基因多于两种时,称为复等位基因(muotiplealleles)。HLA复合体Ⅰ类和Ⅱ类基因位点多为复等位基因。1995年公布的用血清学、MLR和PLT确认的HLA特异性见表6-1。
表6-1HLAⅠ类和Ⅰ类基因特异性总表(1995)
| A | B | C | DR | DQ | DP | D | |
| A1 | B7 | B5102 | Cw1 | DR1 | DQ2 | DPw1 | Dw1 |
| A2 | B703 | B5103 | Cw2 | DR103 | DQ4 | DPw2 | Dw2 |
| A203 | B8 | B52(5) | Cw3 | DR2 | DQ5(1) | DPw3 | Dw3 |
| A210 | B13 | B53 | Cw4 | DR3 | DQ6(1) | DPw4 | Dw4 |
| A3 | B15 | B54(22) | Cw5 | DR4 | DQ793) | DPw5 | Dw5 |
| A11 | B18 | B55(22) | Cw6 | DR7 | DQ8(3) | DPw6 | Dw6 |
| A23(9) | B27 | B56(22) | Cw7 | DR8 | DQ9(3) | Dw7 | |
| A24(9) | B35 | B57(17) | Cw8 | DR9 | Dw8 | ||
| A2403 | B37 | B58(17) | Cw9(w3) | DR10 | Dw9 | ||
| A25(10) | B38(16) | B59 | DR11(5) | Dw10 | |||
| A26(10) | B39(16) | B60(40) | DR12(5) | Dw11(w7) | |||
| A29(19) | B3901 | B61(40) | DR13(6) | Dw12 | |||
| A30(19) | B3902 | B62(15) | DR14(6) | Dw13 | |||
| A31(19) | B40 | B63(15) | DR1403 | Dw14 | |||
| A32(19) | B4005 | B64(14) | DR1404 | Dw15 | |||
| A33(19) | B41 | B65(14) | DR15(2) | Dw16 | |||
| A34(10) | B42 | B67 | DR16(2) | Dw17(w7) | |||
| A36 | B44(12) | B70 | DR17(3) | Dw18(w6) | |||
| A43 | B45(12) | B71(70) | DR18(3) | Dw19(w6) | |||
| A66(10) | B46 | B72(70) | DR51 | Dw20 | |||
| A68(28) | B47 | B73 | DR52 | Dw21 | |||
| A69(28) | B48 | B75(15) | DR53 | Dw22 | |||
| A74(19) | B49(21) | B76(15) | Dw23 | ||||
| B50(21) | B77(15) | Dw24 | |||||
| B51(5) | B7801 | Dw25 | |||||
| Dw26 | |||||||
HLA抗原的命名由世界卫生组织命名委员会确定,每个特异性抗原均以其基因位点的字头附以适当的数字(按抗原被发现或官方认可的顺序)表示。标有w(workshop)的为暂用名,得到认可后将其去掉;1991年决定:新特异性的申报要有明确的DNA顺序,并根据DNA间关系命名,故取消w;现在所保留的w已非当初实验室暂定名的含义,例如保留Cw以示与补体缩写区别,保留Dw和DPw以示其用细胞学方法检测。后面带括弧的表示该特异性由括弧内的特异性分解而来,括弧内为早期确认的抗原,包含多个特异性。表中D抗原不是独立基因位点的编码产物,而是与DR和DQ广泛相关,是用细胞学方法检测的抗原。
表6-1所列特异性是用血清学方法和细胞学方法鉴定出来的,几乎每次会议都命名新的特异性。如此复杂的基因及产物,再加上单倍体共显性遗传的特点,可随机组合成一个巨大的数字;以致在人群中除同卵双胎外,难以找到HLA完全相同者。这充分体现了HLA对免疫调控的个体差异,也为同种器官移植增加了困难。
现在用分子生物学方法可在基因水平上鉴定出更大的HLA多态性,例如HLA-A2的基因有12个变异体(A*0201~A*0212),其差别仅在第19密码子一个碱基的置换。1994年3月WHO命名委员会公布的Ⅰ类和Ⅱ类等位基因为440个,1995年1月又发现了35个新的基因序列,并对以前的报告进行了部分修正。
三、HLA的遗传特点
1.单倍型遗传单倍型(haplotype)是指一条染色体上HLA各位点基因紧密连锁组成的基因单位。人体细胞为二倍体型,两个单倍型分别来自父亲和母亲,共同组成个体的基因型(genotype)。由于一条染色体上HLA各位点的距离非常近,很少发生同源染色体之间的交换,因此新代的HLA以单倍型为单位将遗传信息传给子代。例如父亲的基因型为ab,母亲的为cd,则子代可能有4种基因型,ac,ad,bc,bd,某一个体获得任一基因型的可能性都是1/4。故两个同胞有完全相同或完全不同HLA基因型的可能性都是1/4;一个单倍型相同的可能性是1/2。而子代和亲代总是共有一个相同的单倍型。
2.共显性遗传共显性(co-dominance)是指某位点的等位基因不论是杂合子还是纯合子,均能同等表达,两者的编码产物都可在细胞表面检测到。故每个位点可具有两个抗原,可能相同,也可能不相同;这些抗原组成了个体的表型(phenotype)。多数个体的HLA位点都是杂合子,但当父亲和母亲在某位点上具有相同的等位基因时,其子代的这个位点就成为纯合子。
3.连锁不平衡理论上,一个HLA位点的等位基因与另一个或几个位点的等位基因在某一单倍型出现的频率应等于各自频率的乘积。然而在很多情况下,预期的单倍型频率往往与实际检测的频率相差很大,在不同的地区或不同的人群,某些基因相伴出现的频率特别高,这种现象称为连锁不平衡(linkagedisequilibrium)。HLA基因连锁不平衡的发生机制目前尚不清楚,但已经发现某些疾病的发生与HLA复合体中某些特定的等位基因密切相关;某些连锁不平衡倾向于出现在某些区域、某些人种和某些民族。深入探讨连锁不平衡的发生机制无疑将有助于对某些疾病的诊断和治疗,亦将为人类学研究增添新的内容。
四、HLA的分型技术
HLAⅠ类抗原的DQ、DR用血清学检测法进行分型,因此在方法学上称为血清学鉴定的抗原(serologicallydefinedantigen,SD抗原);DP和D特性需用细胞学方法进行检测,因此称为淋巴细胞鉴定的抗原(lymphocytedefinedantigen,LD抗原)。虽然HLA的基因分型技术发展很快,但目前仍不能完全取代血清学分型法和细胞分型法(详见第二十八章)。